

SARS-CoV-2蛋白互作网络图为老药新用提供新思路
2020-09-27 来源:Nexcelom 点击次数:3942
研究设计思路
研究者们克隆并在人细胞系HEK293T/17中表达了29个SARS-CoV-2蛋白中的26个,然后用亲和纯化/质谱(AP-MS)确定了332个高置信度的SARS-CoV-2-人蛋白互作(PPIs)。从这些PPI中,他们找到了被69种已知药物(29个FDA批准的药物,12个处于临床实验阶段的药物,28个临床前化合物)靶向的66个人蛋白或宿主因子。接下来,来自美国西奈山医院和法国巴斯德研究所的两个团队各自利用Celigo免疫荧光成像分析和qRT-PCR的方法对一部分药物进行了体外实验筛选,发现两类化合物具有病毒抑制能力:1)mRNA转译抑制剂和 2)Sigma1和Sigma2受体的潜在调节剂。对这些靶向宿主因子的化合物进行更深入的研究,以及探索与靶向病毒蛋白的药物的联合使用,可能会成为COVID-19药物开发的新思路。
以下是此篇文章的重点结果和方法解读。
SARS-CoV-2宿主互作蛋白的全面分析
通过AP-MS, 研究者们确定了332个高置信度的SARS-CoV-2-人蛋白互作,并对这些人蛋白的生物学功能和组织表达水平进行了研究(Fig. 1a)。SARS-CoV-2的蛋白参与了一些重要的宿主细胞活动,包括脂蛋白代谢(S),细胞核运输(Nsp7),和核糖核蛋白复合物合成(Nsp8)(Fig. 1b)。在29个人类组织中,相关蛋白在肺中的表达水平最高(Fig. 1c)。在感染过程中,病毒蛋与其相互作的人蛋白的丰度变化有很强的相关性(Fig. 1d)。通过与其他病原体-宿主的蛋白互作网络图相比较,研究者们发现西尼罗病毒(WNV)和结核分支杆菌(Mtb)与SARS-CoV-2拥有最相似的网络分布(Fig. 1e)。值得注意的是,结核分支杆菌也可以感染肺组织。
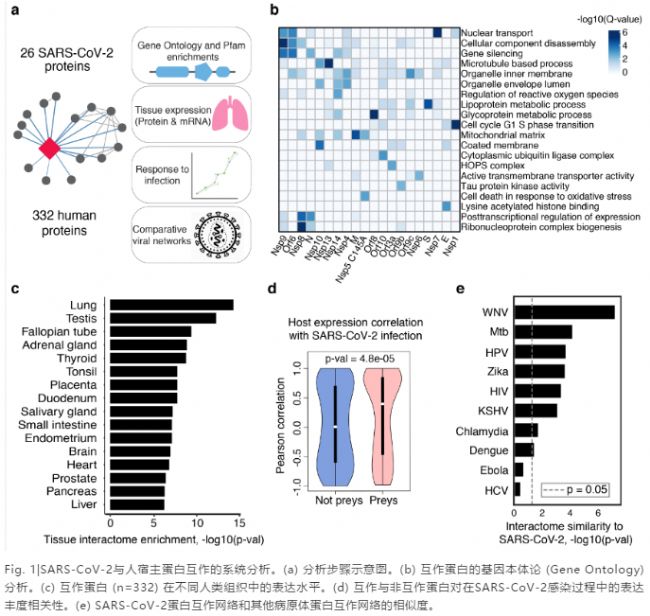
蛋白互作网络图刷新了人们对SARS-CoV-2的认知
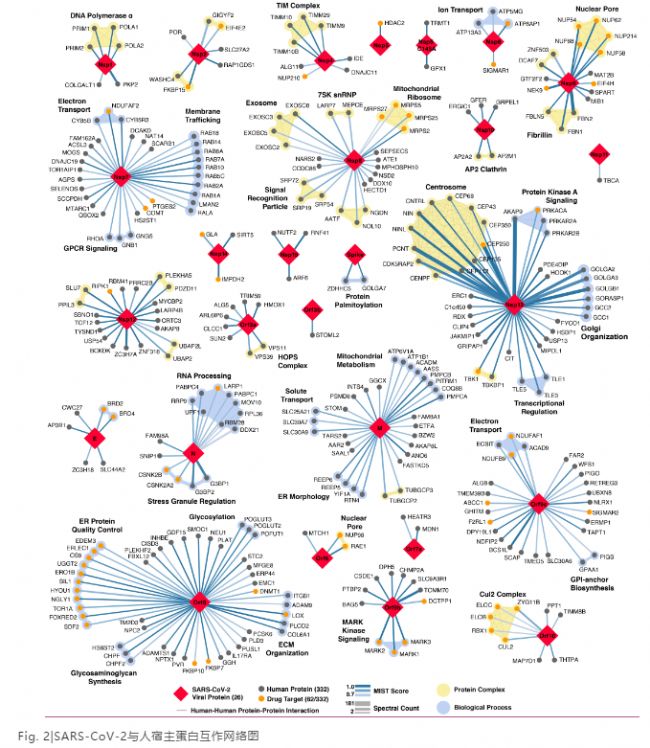
SARS-CoV-2与人蛋白之间复杂的互作网络涉及了多个复合物和生物过程,包括DNA复制,表观遗传和基因表达调控,囊泡运输,脂类修饰,RNA加工和调节,泛素连接酶,信号传导,细胞核运输,细胞骨架,线粒体和细胞外基质(Fig. 2)。大约40%的SARS-CoV-2互作蛋白与内膜或囊泡运输路径有关。这些互作可能在病毒进入宿主细胞时起到辅助作用。
研究还发现SARS-CoV-2与天然免疫信号传导相关蛋白有相互作用。比如Nsp13, Nsp15和Orf9b与IFN路径有关联; Nsp13和Orf9与cNF-κB路径有关联。另外,两个调控抗病毒天然免疫信号传导的E3泛素连接酶,TRIM59和MIB1,分别和Orf3a与Nsp9相结合。
SARS-CoV-2还会影响宿主细胞蛋白的转译过程。病毒的核衣壳可以结合应激颗粒(SG)蛋白G3BP1/2,还有mTOR调控的转译抑制因子LARP1, 蛋白激酶CK2,mRNA降解因子UPF1和MOV10。应激颗粒的形成是最早发生的抗病毒反应之一。对应激颗粒和相关RNA路径进行操控在冠状病毒中很常见。由eIF4A抑制剂促进的G3BP聚集也许是其抗病毒能力的原理。
靶向SARS-CoV-2人源宿主因子的现有药物
借助化学信息学工具,研究者们找到了靶向SARS-CoV-2人源宿主因子的16个批准药物,3个在研药物(临床实验阶段),以及18个临床前候选药物;另外,通过基于靶点和信号通路的文献搜索,他们还发现了13个批准药物,9个临床实验阶段的在研药物(INDs),和10个临床前候选药物。在所有332个高置信度人蛋白靶点中,有63个拥有已知靶向分子,总计69种批准药物/INDs/临床前化合物。
宿主靶向药物的病毒抑制能力
接下来,研究者们对这些药物的抗病毒活性进行了分析。这项工作由来自纽约的西奈山医院和巴黎的巴斯德研究所的两个团队分别完成。西奈山医院团队开发了一种中等通量的基于Celigo免疫荧光成像的方法(检测病毒核蛋白NP),在非洲绿猴肾细胞系Vero E6中对37种化合物进行了SAR-CoV-2感染抑制筛选。巴斯德所团队则通过qRT-PCR检测细胞上清病毒RNA的方法分析了44种药物的病毒抑制能力(Fig. 3a)。两个团队一共对69个药物中的47个进行了检测,另外还有13个靶向SaigmaR1/R2受体和mRNA转译的化合物,以及额外的15个其他方法筛选出的小分子。
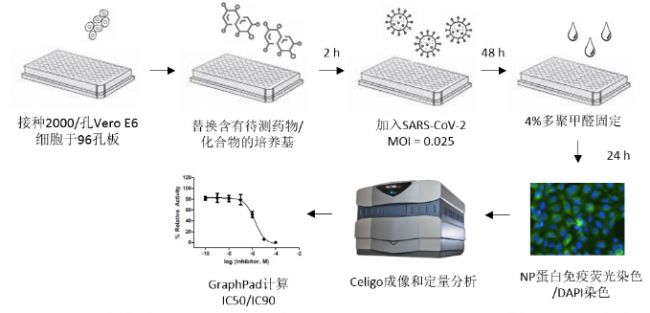
病毒抑制实验方法(西奈山医院)
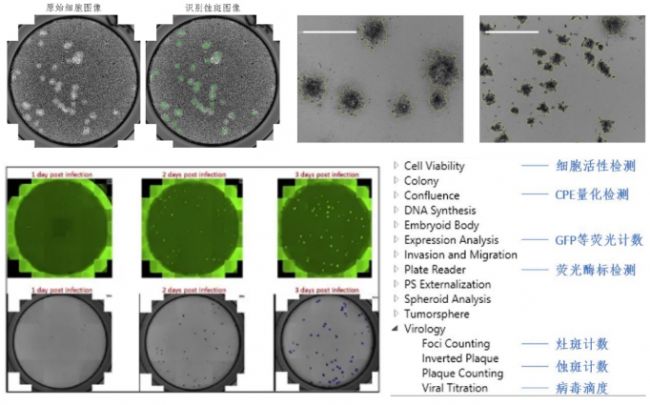
将Vero E6细胞以2000细胞/孔接种于96孔板(由于Vero E6是肾细胞系,因此不排除药物在肺细胞中会产生不同的结果)。病毒感染前2小时,将培养基替换为含有待测药物/化合物的培养基,对照孔加入相应浓度的DMSO。将培养板转移至P3安全等级实验室,加入100 PFU (MOI 0.025)的SARS-CoV-2,于37C培养48小时。感染完成后,去掉上清,向板中加入4%多聚甲醛固定细胞。24小时后,将培养板移出P3实验室。接下来,对细胞中的病毒核蛋白(NP)进行免疫荧光染色(SARS-CoV 抗血清,1:10000),并用DAPI染细胞核。Celigo成像细胞分析仪(Nexcelom Bioscience)被用于对感染的细胞(AF488)和总细胞(DAPI)进行荧光成像和定量分析。细胞中绿色荧光(NP)的强度反映了病毒感染的程度。感染率百分比可以通过((感染细胞数/细胞总数) – 背景) x 100%计算出。DMSO对照组的感染率被设置为100%以完成归一化。IC50和IC90值通过将Celigo的数据导出至GraphPad分析得出。
对于挑选出的药物,研究者们用被感染细胞的上清进行了病毒滴度检测并计算出TCID50。具体步骤为:SARS-CoV-2感染48小时后收集细胞上清,冻存于-80 ˚C直至使用。将Vero E6细胞以20000细胞/孔接种于96孔板。第二天,加入10倍梯度稀释(10^1 到10^6)的含SARS-CoV-2上清的培养基。五天后,将细胞用结晶紫染色,观察细胞病变(CPE)。用Reed&Muench方法计算TCID50/mL。
病毒抑制实验的前几个步骤和西奈山医院团队的方法基本一致(不过每孔加入SARS-CoV-2的MOI为0.1)。病毒感染48小时后,收集细胞上清用于RNA提取和qRT-PCR。根据已知病毒滴度做出的RNA标准曲线,计算出上清中病毒基因对应的PFU数值。
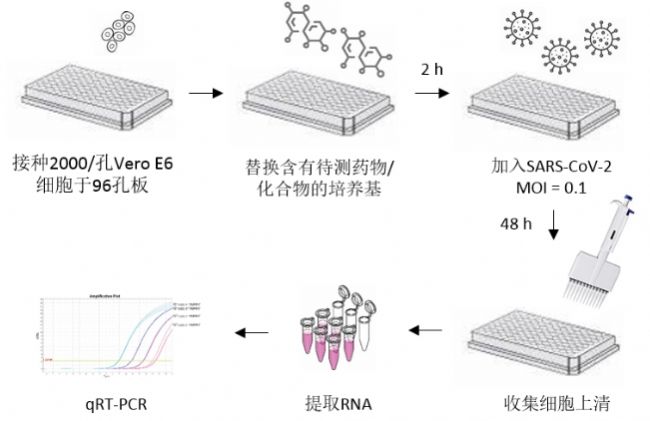
病毒滴度检测(巴斯德所)
病毒的滴度是通过蚀斑实验来检测的。将Vero E6细胞以7.5x10^4/孔的密度接种于24孔板。第二天,加入10倍稀释的含病毒培养基,于37˚C孵育1小时。随后,加入0.005%琼脂糖形成半固体覆盖层。于37˚C孵育3天后用4%多聚甲醛固定和结晶紫染色,肉眼观察和计数每孔蚀斑数量(plaque assay)。
西奈山医院团队用于检测药物抑制病毒能力的仪器为Celigo高通量成像细胞分析仪。该仪器配备有1个明场和4个荧光通道,能对生长在微孔板中的细胞进行高速全孔高清成像(检测一块96孔板的病毒抑制实验 < 15分钟),并用内置软件识别每一个细胞,完成多重参数(包括细胞数量、大小、平滑度、平均荧光强度、总荧光强度、不同类型细胞百分比等)的定量分析。
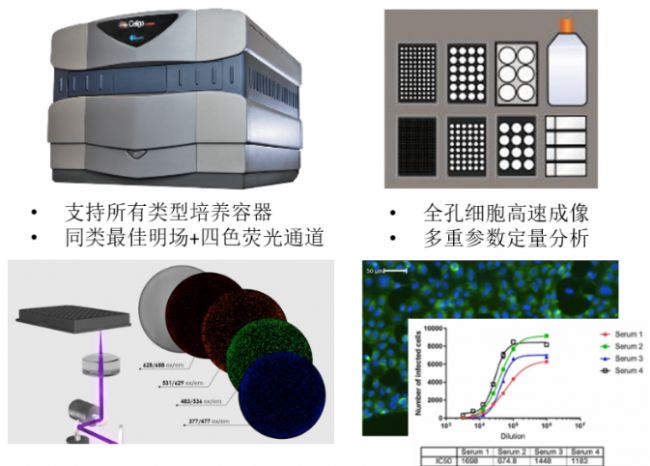
病毒抑制/抗体中和的常用实验方法有很多种,包括如蚀斑减少中和实验(PRNT),病毒灶减少中和实验(FRNT)和基于ELISA的微中和实验等。与这些方法相比,基于自动荧光成像的方法节省人力和时间,减少了人为误差,通量高,灵敏度高,动态范围广,所需病毒量少(MOI低),非常适合抗病毒药物的高通量筛选。
值得注意的是法国巴斯德所团队采用的方法更为传统。qRT-PCR具有很高的灵敏度,因此也更容易产生交叉污染,对实验者的操作有很高的要求。另外,qRT-PCR反映的是RNA总量,不能直接体现病毒的感染复制能力。
用于病毒滴度检测的蚀斑实验通常只能肉眼人工计数,速度慢,通量低,且无法避免人为误差。Celigo可以对蚀斑实验的孔板进行全孔成像,软件自动识别蚀斑,得到每孔蚀斑数量、大小、平滑度等多个参数。Celigo也能检测HRP或荧光染色的病毒灶斑,甚至分析单个细胞水平的病毒感染率。
细胞毒性实验
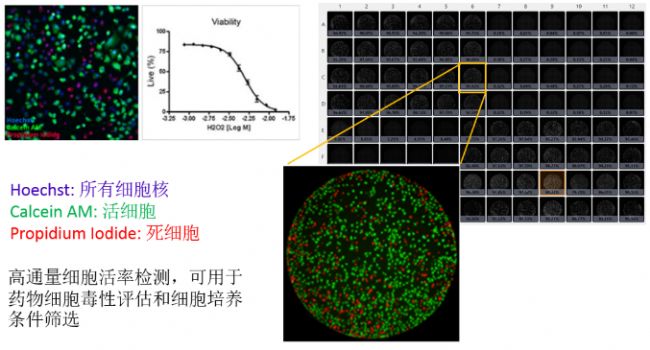
西奈山医院团队用MTT方法对有和没有感染SARS-CoV-2的Vero E6细胞进行了药物浓度梯度的毒性实验。巴斯德所团队用阿尔玛蓝试剂检测药物对细胞存活率的影响。细胞存活率百分比通过基于健康细胞(100%)和乙醇处理的细胞(0%)做出的标准曲线计算得出。
药物安全性评估是药物筛选中非常重要的一个环节。常用的检测细胞毒性的方法大多基于细胞的酶活性和能量代谢,比如本文作者使用的MTT和阿尔玛蓝,还有CCK-8, CellTiterGlo等。然而越来越多的研究显示这类方法得出的药物对细胞活性的影响与实际有偏差,尤其是能直接影响细胞酶活性和能量代谢的药物。
基于荧光染色的细胞计数是目前公认最准确的细胞活性检测方法。不同荧光染料的组合包括AO/PI, Calcein AM/PI, PI/Hoechst等。对于西奈山团队的实验设置,在病毒感染和药物处理过程中加入PI,免疫荧光和DAPI染色后即可在同一块板上检测药物毒性。Celigo可以快速准确地得出整板细胞数和存活率,非常适合于高通量药物细胞毒性评估。
mRNA转译和Sigma受体抑制剂的SARS-CoV-2抑制作用
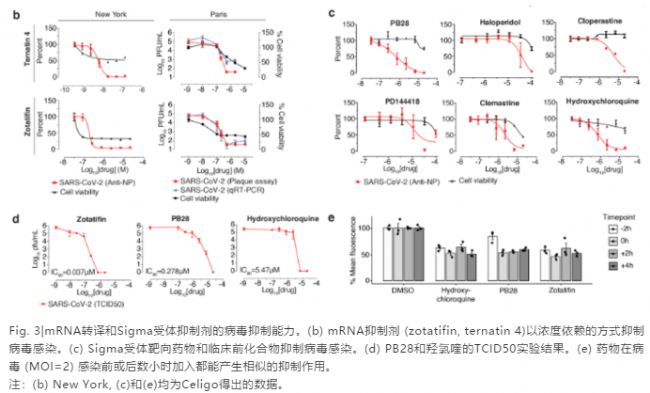
两个团队的实验结果显示有两类化合物可以抑制病毒感染:mRNA转译抑制剂(zotatifin, ternatin-4和PS3061; Fig. 3b, Extended Data Fig. 9)和Sigma1与2受体的抑制剂(包括haloperidol, PB28, PD-144418和正处于临床实验阶段的羟氯喹)。其他的Sigma1和Sigma2受体抑制剂clemastine, cloperastine和孕酮(Fig 3c, Extended Data Fig. 9)也具有病毒抑制能力。Fig. 3d展示的是用TCID50方法做出的部分药物病毒抑制曲线。值得注意的是,该结果显示PB28(IC90 = 280 nM)比羟氯喹的效力强~20倍。
西奈山团队也用Celigo荧光成像法检测了病毒感染前后不同时间加药对病毒感染(荧光强度)的影响。该实验用较高滴度(MOI=2)的病毒对细胞进行单轮感染(8小时)。结果显示羟氯喹、PB28和zotatifin不论在感染前还是感染后加入,对病毒感染的抑制能力是相似的(Fig. 3e),说明这些药物是在病毒复制的过程中发挥作用的。
研究者们相信,靶向Sigma1和2受体的小分子以不同于转译抑制的路径抑制病毒感染,比如通过调控细胞应激反应。值得关注的是,一些靶向Sigma受体的化合物,比如clemastine, cloperastine和孕酮,都是经FDA批准上市多年的药物。另外,Sigma受体靶向化合物在有明显病毒抑制能力的同时具有较小的细胞毒性(Fig. 3b和c)。
Sigma1和2受体属于胞内侣伴蛋白,在中枢神经系统高度表达。相对于Sigma1受体,人们对Sigma2受体的了解较少。Sigma1受体可以调控电压依赖的离子通道,因而影响神经元的激活和突触活动,在突触可塑性,学习和记忆中都起到重要作用。此外,有证据表明Sigma-1受体和帕金森以及重度抑郁症有关联,因此成为这类疾病的热门靶点。
讨论与总结
本次研究获得的化学-蛋白质组学分析结果不仅提供了以SARS-CoV-2的人互作蛋白为靶点的潜在化合物信息,也揭示了这些化合物可能的作用机制。mRNA转译抑制剂显示出较强的病毒抑制作用(有效浓度在10 – 100 nM之间),使得这类化合物成为极具吸引力的潜在药物。虽然靶向Sigma1和2受体化合物的作用机制还不清楚,但是它们对病毒感染的控制能力值得关注。PB28的高效力(280 nM IC90)和高特异性使其拥有很高的成药潜力。还有许多已批准的Sigma靶向药物拥有老药新用的潜能,值得评估其对SARS-CoV-2的抑制能力。
作为抗病毒疗法,宿主靶向的干预方式可以避免因病毒变异而产生的药物抗性,也为开发广谱抗病毒疗法从而应对未来其他病毒的爆发提供了可能性。另外,宿主靶向药物可以和病毒靶向药物(比如瑞德西韦)组成联合疗法,以达到更好的病毒抑制效果。更重要的是,本次研究所展示的工作流程代表了药物发现(不局限于广谱抗病毒疗法,也适用于其他疾病疗法的开发)的一种新方法,也是国际间多学科合作促进科学发展的一次完美体现。