

关于人类多选择知觉选择中的神经动力学决策变量的研究
2024-12-24 来源:本站 点击次数:37901研究背景
支持二元知觉决策及其转化为行动的神经动力学得到了很好的研究,但现实世界的决策通常提供两种以上的反应选择。与决策相关的证据积累如何动态地影响人类的多种行为表征这个问题还仍需解决。
与二元选择方案相比,多重选择方案所需的高度保守性表明一种机制可以通过抑制基线活动来补偿存在多项选择时增加的不确定性。这里,我们在四项和两项选择的感知决策中使用皮质脊髓兴奋性跟踪动作表征,并使用顺序采样框架对其进行建模。
本实验旨在使用人类行动表征的动态变化证明泄漏竞争累加器模型做出的预测可以适应多种选择(即,减少基线活动以补偿增加的不确定性)。
02研究方法
2.1、被试
我们招募了13名被试(5名男性),平均年龄为26.23岁(SD=7.67)。因为由于不可避免的原因,许多TMS试验必须被放弃,每个被试完成两到四次治疗(每次持续1-3小时),平均完成4166次试验。
2.2、实验流程
被试完成了一项颜色辨别任务。在每次试验中,都会出现一组彩色像素。数组由四种不同的颜色组成:绿色(RGB [0.6 0])、红色(RGB[0.8 0 0])、黄色(RGB [0.92 0.74 0])、蓝色(RGB [0.120.12 0.61])。
被试被指示使用四种响应中的相应一种,尽快并尽可能准确地指出哪种颜色最普遍按钮(见图 1)。使用 MATLAB生成刺激,并在 Display++ LCD 监视器(剑桥研究系统有限公司;显示尺寸:41 cm × 30 cm) 以 100 Hz 的刷新率和 1240 × 786 像素的分辨率运行。被试坐在离屏幕大约 100 厘米的地方。刺激阵列对着 6 × 6 度的视角。每个彩色像素跨越 2 × 2 屏幕像素,产生 145 × 145 彩色像素阵列。
被试每只手拿着两个通过 16 位 A/D 卡(National Instruments X-seriesPCIe-6323,采样率 100,000 Hz)接口的数字响应按钮,一个在拇指和食指之间(捏),一个靠在手掌上,附在到一个塑料圆柱体(掌握;Hadaret al., 2012)。捏收缩第一背侧骨间 (FDI) 肌肉,同时抓住圆柱体激活小指外展肌 (ADM)。每种颜色都映射到四个响应按钮之一。颜色反应映射在被试之间随机进行。试验包括提示(500 毫秒)、彩色刺激阵列(2000 毫秒或直到响应)和 ISI(最小 500 毫秒)。该实验包含两个和四个选择试验。此外,作为阳性对照,我们在 2 × 2 设计中通过我们对选择数量的操纵跨越了难度操纵。每个试验开始时的提示通过呈现代表可能的选择的两个或四个彩色方块来告知被试给定的试验是两项选择还是四项选择试验。三分之一的试验是四项选择,三分之一是两项选择“在”手,即两种可能的反应是在一只手上(左捏-左抓,右捏-右抓),三分之一的试验是“在”手之间的两个选择,即来自手的同源反应双手都可以(左捏-右捏,左抓-右抓)。试验顺序是随机的。在第一次会面中,被试完成了 150 次练习试验以熟悉任务。然后,被试在每次会面上完成 5 到 6 个Block,每个block包含 168 次试验(加上额外的试验以调节 TMS 频率;见下文)。
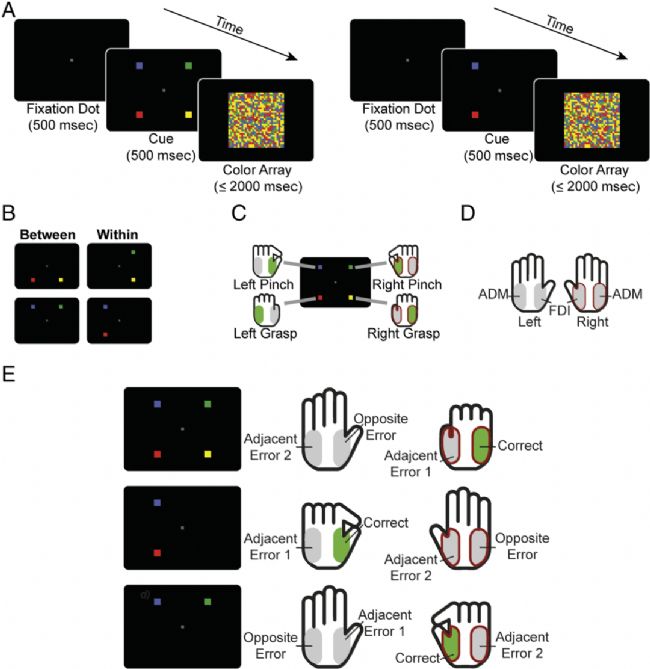
图1. 试验程序。(A) 颜色辨别任务。被试必须指出哪种预制颜色在颜色阵列上最普遍。左:四选试;显示了所有可能的线索。右:二选一试验;只显示了两个可能的线索。(B) 所有可能的二选一组合。在“内”试验中,两种可能的反应都在同一只手上,而在“之间”试验中,两种可能的反应在不同的手上,但使用相同的反应(捏/抓)。(C)每个提示/颜色都与特定反应相关;右上角提示(此处:绿色)与右手“捏”响应相关,左上提示(此处:蓝色)与左手“捏”响应相关,右下提示(此处:黄色)与右手“抓住”反应,左下方提示(此处:红色)与左手“抓住”反应相关联。颜色-响应映射在被试之间随机化,而提示位置-响应映射保持不变。(D) 每只手都执行捏和抓握反应,分别招募 FDI 和 ADM 肌肉。与正确反应相关的肌肉显示为绿色,而其他肌肉显示为灰色(参见 C 和 E)。MEP 仅从右手肌肉记录(由深红色边框表示)。(E) 响应分类。在每个试验中,右手边的 MEP 与两个响应类别相关。由于正确响应的位置是随机变化的,因此所有类别的激活都在试验中进行了探索。顶部:四项选择试验,其中刺激阵列(未显示)指示正确的抓握反应。同一只手上的错误响应标记为“相邻错误 1”,而另一边的响应标记为“相邻错误 2”(与正确响应同源)和“相反错误”(与正确响应非同源)。中间:两个选择“内”试验,这里涉及左手反应之间的选择。如果左捏反应是正确的,不正确但有提示的反应(这里是左抓握)被归类为“相邻错误 1”,另一方面,与正确反应同源的反应被标记为“相邻错误 2”,相反的响应被标记为“相邻错误 2” “相反的错误”。底部:两个选择“之间”的试验,涉及在左捏或右捏反应中进行选择。在这里,右捏是正确的,而被提示但不正确的左捏是“相邻错误 1”。与正确响应在同一只手上的错误响应标记为“相邻错误 2”,而解剖学上相反的响应是称为“相反的错误
2.3、实验刺激与数据采集
使用表面肌电信号记录刺激和记录肌肉活动,通过13位A/D Biometrics Datalink系统(版本7.5,Biometrics Ltd.)以1000 Hz采样。和带通滤波(20-450 Hz)。表面Ag/AgCl电极(22 mm×28 mm,部件号。生物识别有限公司SX230FW)。放在每只手的FDI和ADM之上。参比电极被放置在距离四个有源电极中的每一个大约2 cm的地方。右侧ADM和FDI的肌电信号也被传递给分别位于参与者左侧和右侧的扬声器,噪音通知参与者,他们的肌肉在两次反应之间没有完全放松。使用Magstim Rapid2双相刺激器(The Magstim Co.Ltd.)进行单脉冲TMS。为了诱导右手ADM和FDI的MEP,在左侧运动皮质上方的头皮上放置了一个70 mm的八字形线圈(每个线圈的外壳直径约为90 mm)。每个参与者的确切位置和刺激强度都被单独调整,并被设定为静息运动阈值的大约110%。静息运动阈值被定义为在50%的右手直接刺激和右手ADM刺激中诱发峰间振幅约为50μV的MEP的最小强度。使用神经导航系统(荷兰Ant Neuro的Visor 2)连续跟踪和维持TMS的位置。在每种情况下57%的试验中都计划了TMS脉冲。为了确保TMS脉冲在基线间隔和RT过程中的良好分布,TMS试验被分成四个频率相等的时间段,它们位于−相对于刺激开始的200毫秒和700毫秒之间(即相对于线索开始在300毫秒到1200毫秒之间)。在给定箱内,随机抽取每个试验的确切刺激时间。因为实验遵循单脉冲TMS方案,所以脉冲需要以0.2 Hz的最大频率出现。如果碰巧计划的脉冲间间隔(IPI)小于5000毫秒,则调整定时。对于4-5000毫秒的IPIs,增加了发作间隔,将脉冲频率降低到0.2 Hz。对于小于4000毫秒的IPIS,计划中的试验被下一个计划中的无刺激试验所取代。如果没有剩余的无刺激试验,则产生随机无刺激试验以增加IPI。平均而言,这种方法每期增加434次试验,平均每期试验1354次。如果已经检测到响应,则不传送计划的脉冲。
2.4、数据处理与分析
整理和平滑对于每个正确/错误类别和每种情况,在正确试验中记录的MEP被跨参与者和会话汇集在一起。因此,我们对每个会话和参与者的刺激时间进行了标准化,将其表示为中位肌电图RT的百分比(在无刺激试验中)。汇集的MEP波幅在时间上进行了排序,并与刺激和反应保持一致。
统计分析我们报告所有配对检验的双尾值。行为数据条件(即RTS、EMG RTS和误差)之间的差异分别使用方差分析和带有Logistic链接函数的广义线性混合模型来推断。使用MATLAB fitglme命令实现广义线性混合模型,所有感兴趣的影响(例如,难度、响应选择的数量及其相互作用)都嵌套在参与者和会话中,并作为随机效果包含在模型规范中。对于TMS数据,基于之前对非人类灵长类动物的研究,我们预计当参与者在四个选项中选择四个而不是两个时,基线MEP振幅会降低。为了检验这一点,我们(为每个参与者)记录了在线索刺激间隔期间记录的与线索反应相关的所有MEP的幅度(即,对于四项选择试验,所有MEP都在基线期间,而对于两项选择试验,只有MEP被归类为“正确”或“相邻错误1”)。在对这些分数进行平移和平方根变换以实现常态之后,使用MATLAB fitlme命令使用线性混合模型(对参与者具有随机效应)来测试条件间的差异(与对多个试验得出的平均值进行的传统t检验相比,该命令更好地利用了来自所有贡献分数的信息)。这一方法也被应用于我们的建模中出现的一个不可预见的预测,即相反的错误和其他(即相邻的)错误类别之间的后期累积剖面的差异。我们还预计MEP信号会受到任务难度的影响,“容易”的测试比“难”的测试积累得更多。在具有多个累加器的序贯抽样模型下,累积率的差异是行为的关键决定因素。因此,我们将重点放在试验中,在这些试验中,一只手的两个MEP可以用来构建一个信号,测量“正确的”和“相邻的错误1”反应之间的差异。对于四个选项和两个选项的手内试验,我们用一条直线拟合了从这种差异得出的MEP信号。我们称这条线的坡度为相对论坡度。我们对“容易”和“难”试验都这样做,然后从与“容易”试验相关的斜率中减去“难”试验的相对化斜率。这些估计是针对刺激锁定数据(50%至90%的中位肌电图RT)和反应锁定数据(−50%至−10%的中位肌电图RT)做出的。我们使用1999次迭代的非参数置换检验来产生新的重采样的“容易”和“困难”条件集(没有替换),并计算它们之间的相对化斜率差。在每项测试中,将原始差异与结果差异的零分布进行比较。
03实验结果
在所有 4320 个配对决策的 90% 中,至少有一个成对成员投资于捕食或猎物防御 ( X , Y > 0)。检查投资选择的频率分布(图 2A 和 B)表明被试确实进行了投资,投资是分散的,并且猎物的投资比掠夺者的投资更多。从什么标准理性的经济理论所预测的(偏离7,13,34),然而,与会者还过度投资(X,Ÿ ≥7),理论上不应该发生。此外,虽然在 TBS 治疗中猎物的投资选择分布相似(图 2A),掠夺者投资确实不同,尤其是在“不攻击”决策的频率上 ( X = 0)(图 2B)。
图 2A 和 B中看到的模式在总体投资和投资频率的 GLMM 分析中得到证实。猎物比捕食者投入更多(F = 48.025,P = 0.001),当 rIFG 上调而不是下调时,这种差异有所增加,但没有显着增加(图 2C;角色 × 治疗,F = 3.629,P = 0.057 )。攻击频率出现了更强大的效果(图 2D)。对于频率,角色×治疗交互作用显着(F = 4.417,P = 0.036)。当他们的 rIFG 相对于假治疗(F = 4.490,P = 0.034)和 rIFG 上调(F = 5.341,P = 0.025)下调时,捕食者会更频繁地投资。
相对于假治疗,下调的 rIFG 产生更多的攻击(根据图 2D)、更多的竞争成功(图 3A;F = 5.451,P = 0.020)和更高的个人收入(图 3B;F = 11.189,P = 0.001) )。事实上,当 rIFG 被下调而不是假处理时,攻击预测了个人收益(图 3C;β = 1.460,t = 3.750,P = 0.001; 攻击 × 治疗,F = 14.059,P = 0.001)。
上调 rIFG 不会导致更多攻击(根据图 2D)或更大的竞争成功(图 3A;F = 1.976,P = 0.160)。尽管上调 rIFG(与假治疗相比)确实会导致更高的个人收入(图 3B,F = 9.992,P = 0.002),但收益并未通过攻击频率预测(图 3C;β = 0.445,t = 1.274,P = 0.213; 攻击 × 治疗,F = 2.100,P = 0.148)。
上调的 rIFG 会导致捕食者攻击更多(图 4A;处理 × Δ-Prey,F = 6.188,P = 0.010)。换句话说,当猎物降低防御时,具有上调 rIFG(相对于假动作)的捕食者更有可能进行投资。重要的是,这种对 Δ-Prey 的差异跟踪也解释了当捕食者 rIFG 上调时更高的收益(图 4B,相对于假处理:处理 × Δ-Prey,F = 4.081,P = 0.044)。因此,随着 rIFG 的上调,捕食者的攻击更多地依赖于降低的猎物防御,因此比接受假治疗时赚得更多。
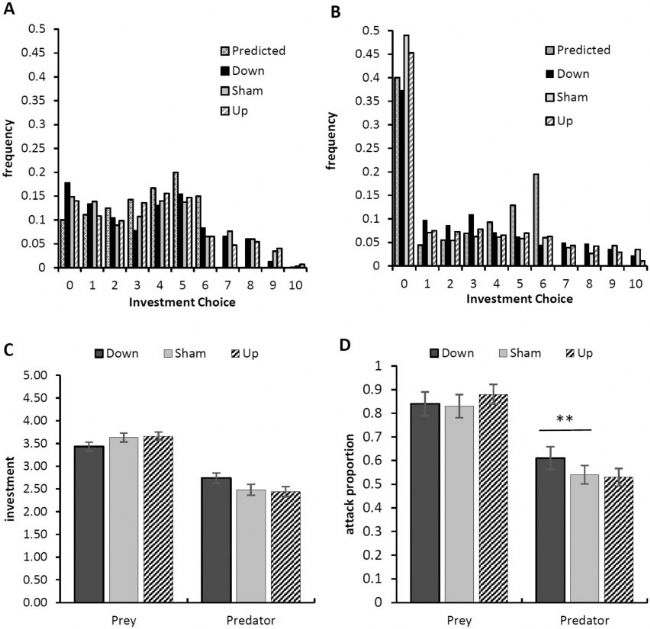 图2. 被试作为捕食者和猎物的投资决策。(A) 按猎物划分的投资决策分布,按处理细分,并与基于标准理性选择理论的预测值进行比较(博弈论预计没有猎物投资≥7)。(B) 掠夺者做出的投资决策分布,按处理细分,并与基于标准理性选择理论的预测值进行比较(博弈论预计没有投资≥7)。(C) 投资作为治疗和角色的函数(显示平均值 ± SE)。(D) 攻击 ( X , Y > 0) 作为治疗和作用的函数(范围 0.0-1.0 对应于 0-40 次攻击决定;显示平均值 ± SE;* P ≤ 0.05;** P ≤ 0.005)
图2. 被试作为捕食者和猎物的投资决策。(A) 按猎物划分的投资决策分布,按处理细分,并与基于标准理性选择理论的预测值进行比较(博弈论预计没有猎物投资≥7)。(B) 掠夺者做出的投资决策分布,按处理细分,并与基于标准理性选择理论的预测值进行比较(博弈论预计没有投资≥7)。(C) 投资作为治疗和角色的函数(显示平均值 ± SE)。(D) 攻击 ( X , Y > 0) 作为治疗和作用的函数(范围 0.0-1.0 对应于 0-40 次攻击决定;显示平均值 ± SE;* P ≤ 0.05;** P ≤ 0.005)
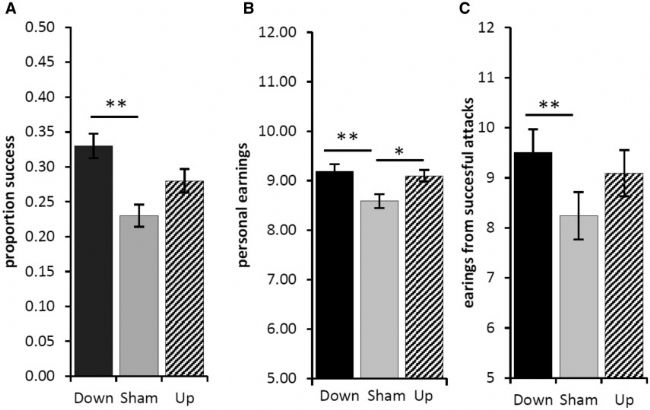 图3. 捕食者高射策略的出现和有效性。(A) rIFG 处理预测捕食者的竞争成功(范围 0.0-1.0,与 40 次可能的胜利成正比;显示平均值 ± SE;* P ≤ 0.05;** P ≤ 0.005)。(B) rIFG 处理预测捕食者收入(范围 0-19 欧元;显示平均值 ± 2SE;* P ≤ 0.05;** P ≤ 0.005)。(C) 作为 rIFG 处理函数的捕食者收益(范围 0-19 欧元;显示平均值 ± 2SE;* P ≤ 0.05;** P ≤ 0.005)
图3. 捕食者高射策略的出现和有效性。(A) rIFG 处理预测捕食者的竞争成功(范围 0.0-1.0,与 40 次可能的胜利成正比;显示平均值 ± SE;* P ≤ 0.05;** P ≤ 0.005)。(B) rIFG 处理预测捕食者收入(范围 0-19 欧元;显示平均值 ± 2SE;* P ≤ 0.05;** P ≤ 0.005)。(C) 作为 rIFG 处理函数的捕食者收益(范围 0-19 欧元;显示平均值 ± 2SE;* P ≤ 0.05;** P ≤ 0.005)
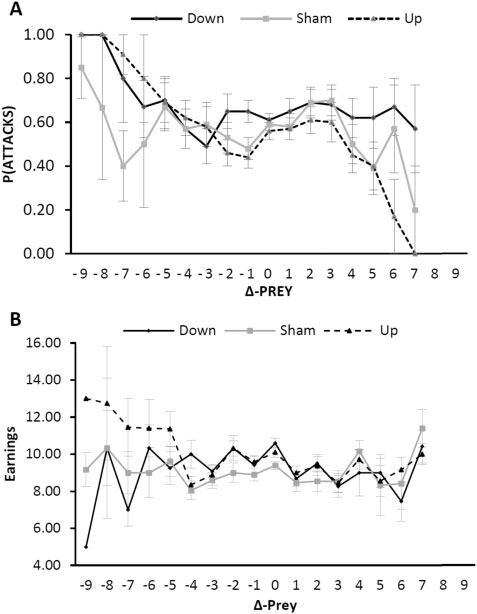
图4. 捕食者跟踪攻击策略的出现和有效性。(A) rIFG 处理和 Δ-Prey 预测捕食者攻击(范围 0.0-1.0,与 40 次可能的胜利成正比;-10、-9、+10 的 Δ-Prey 值由于低 (<2) 观察而未显示;已显示平均值±SE)。(B) rIFG 处理和 Δ-Prey 预测捕食者收入(范围 0-19 欧元;-10、-9、+10 的 Δ-Prey 值由于低 (<2) 观察而未显示;显示平均值 ± SE )
04结论
有效竞争不需要计算思维,尤其是当目标是生存时。增强的前额叶控制使个人在不牺牲竞争效率的情况下显得不那么具有攻击性——它为人类捕食者提供了戴着天鹅绒手套的铁拳。
05参考文献及DOI号
De Dreu, C. K., Kret, M. E., & Sligte, I. G. (2016). Modulating prefrontal control in humans reveals distinct pathways to competitive success and collective waste. Social Cognitive and Affective Neuroscience, 11(8), 1236-1244.
Doi: 10.1093/scan/nsw045