

PLA原位技术在RAD52维持复制叉稳态分子新机制研究中的应用
2026-07-14 来源:本站 点击次数:22复制压力诱导的复制叉停滞与结构紊乱,是驱动基因组不稳定、诱发肿瘤发生以及导致肿瘤靶向耐药的核心分子诱因。2025 年发表于《Nature》的研究《The RAD52 double-ring remodels replication forks restricting fork reversal》,阐明了人源 RAD52 蛋白的全新功能机制:RAD52 可通过独特的双环结构重塑停滞复制叉,调控复制叉稳态,限制 SMARCAL1 介导的异常叉重塑过程,最终维持基因组稳定。该研究中,PLA 邻位连接技术凭借原位检测、高特异性、可精确定量的核心优势,提供了关键的细胞原位实验证据,突破了传统技术无法解析复制叉微环境内分子互作的技术瓶颈。

复制叉周边的分子互作具有瞬时性、弱结合性、空间依赖性的特点,常规实验技术难以实现精准解析:
- Co-IP 技术需破碎细胞提取蛋白,仅能验证体外水平的蛋白结合,完全丢失了细胞内的原位空间信息与时序动态特征;
- 普通免疫荧光(IF)共定位仅能判断两种分子的空间位置重叠,无法区分随机共定位与功能性的相互作用;
- Western Blot 仅能反映细胞内蛋白的总表达量,无法捕捉复制应激状态下,蛋白与复制叉结合的动态变化差异。
受限于上述技术局限,RAD52 不同结构域的功能差异、RAD52 与 SMARCAL1 在复制叉处的竞争调控机制,长期缺乏直接的细胞原位证据,而 PLA 技术恰好补齐了原位定量、特异性互作检测的技术空白。
二、PLA 技术支撑的核心实验发现1、验证 RAD52 多结构域协同介导复制叉原位结合
检测方案:研究使用 IdU 标记细胞内停滞复制叉的新生 ssDNA,结合 PLA 技术特异性检测 RAD52 与复制叉 ssDNA 的原位结合情况,同步对比野生型 RAD52、内侧结合缺陷突变体(IBD)、外侧结合缺陷突变体(OBD)三者的复制叉结合能力差异。
实验结果:在复制压力诱导条件下,野生型 RAD52 可高效富集于停滞复制叉位点,细胞核内的 PLA 特异性荧光信号显著增多;IBD 突变体的复制叉结合信号出现一定程度减弱,而 OBD 突变体的复制叉结合能力大幅下降,PLA 信号衰减最为显著(Fig.1i)。定量统计显示,不同组别细胞核内的 PLA 斑点数量存在显著差异,明确了结构域缺陷对蛋白结合能力的影响。
研究价值:该实验在活细胞原位层面证实,RAD52 对停滞复制叉的锚定依赖多个结构域的协同作用,其中 OBD 外侧结合位点是 RAD52 锚定复制叉的核心功能区域,为 RAD52 双环结构在细胞内的生理功能提供了直观的可视化原位证据。
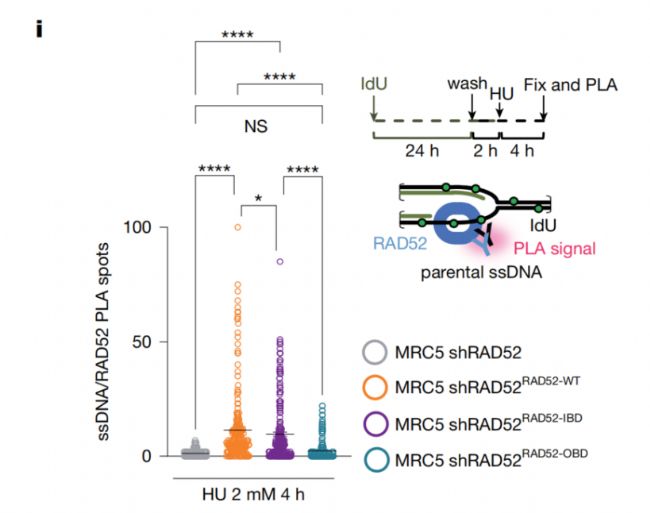 图:PLA 定量分析以单个细胞核内的 PLA 斑点数为统计指标,统计学检验采用 Kruskal–Wallis 检验,结果显示野生型与 OBD 突变体组差异极显著(***P < 0.0001),野生型与 IBD 突变体组差异显著(P = 0.0121)。
图:PLA 定量分析以单个细胞核内的 PLA 斑点数为统计指标,统计学检验采用 Kruskal–Wallis 检验,结果显示野生型与 OBD 突变体组差异极显著(***P < 0.0001),野生型与 IBD 突变体组差异显著(P = 0.0121)。
检测方案:研究采用 PLA 技术定量检测 SMARCAL1 与复制叉 ssDNA 的原位结合水平,结合 RAD52 敲低、野生型 RAD52 及不同突变体回补实验,验证二者在复制叉位点的原位竞争调控关系。
实验结果:敲低 RAD52 后,SMARCAL1 大量富集于停滞复制叉,PLA 原位结合信号显著上调;回补野生型 RAD52 可有效排斥 SMARCAL1,降低其在复制叉处的结合水平;而回补 IBD、OBD 突变体均无法恢复 RAD52 的竞争调控功能,其中 OBD 突变体的功能缺陷最为明显(Fig.4f–k)。
研究同步搭配质谱实验验证二者的结合竞争关系,PLA 成像结果与质谱结论形成交叉验证。
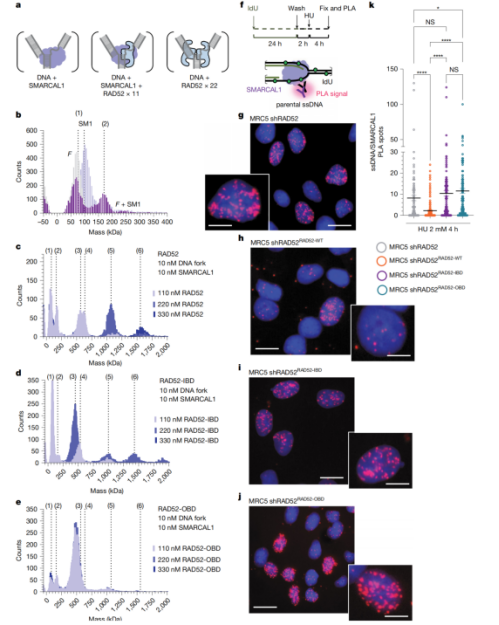
图 4 补充说明:RAD52 与 SMARCAL1 的复制叉竞争结合依赖外侧 DNA 结合位点的完整性。a 为质谱实验的预期分子种类示意图;b-e 为不同 RAD52 变体条件下,RAD52、复制叉、SMARCAL1 形成复合物的质谱检测结果,分别对应无 RAD52、野生型 RAD52、RAD52-IBD 突变体、RAD52-OBD 突变体四组条件;f 为 PLA 检测 SMARCAL1 与亲代 ssDNA 互作的实验流程示意图;g-j 为不同处理组的 PLA 代表性成像图,分别对应 RAD52 敲低组、回补野生型 RAD52 组、回补 RAD52-IBD 组、回补 RAD52-OBD 组;k 为各组细胞核内 PLA 斑点数的定量统计结果,统计学差异采用 Kruskal–Wallis 检验。
研究价值:这一结果在细胞原位层面直接证实,RAD52 通过 OBD 结构域锚定停滞复制叉,以空间占位的方式竞争性排斥 SMARCAL1,进而限制复制叉的异常重塑、维持复制叉结构稳定,完整阐明了 RAD52 双环结构介导的全新复制叉保护机制。
三、研究总结这项发表于《Nature》的研究,依托 PLA 邻位连接技术的原位检测优势,完整阐明了 RAD52 维持复制叉稳态的核心分子机制:RAD52 依靠自身双环结构锚定停滞复制叉,通过 OBD 结构域竞争性调控 SMARCAL1 的结合,从而限制复制叉异常重塑、维持基因组稳态。
与传统蛋白互作检测手段相比,PLA 技术解决了复制叉微环境中瞬时弱互作、原位信号难捕捉的科研痛点,是 DNA 复制应激、基因组稳定性调控、蛋白竞争结合机制等高分研究的核心技术工具,可广泛应用于蛋白 - 蛋白、蛋白 - DNA 的原位互作验证与分子机制解析。
参考文献Honda, M., Razzaghi, M., Gaur, P. et al. The RAD52 double-ring remodels replication forks restricting fork reversal. Nature 641, 512–519 (2025)
蛋白互作PLA邻位连接技术服务哪里有?
乐备实(LabEx)提供专业的 PLA 空间蛋白互作技术服务,拥有成熟的实验平台与丰富的项目执行经验,可覆盖蛋白 - 蛋白互作检测、蛋白翻译后修饰分析等多类需求,支持细胞、组织等多种样本类型的原位检测与可视化分析,为科研、药物研发等场景提供高质量、标准化的技术服务,助力高效完成空间蛋白互作相关研究。
| 货号 | 产品名 |
| LX-PLA | PLA可视化蛋白互作服务 |
乐备实(上海优宁维生物科技股份有限公司旗下全资子公司),是国内专注于提供高质量蛋白检测以及组学分析服务的实验服务专家,自2018年成立以来,乐备实不断寻求突破,公司的服务技术平台已扩展到单细胞测序、空间多组学、流式检测、超敏电化学发光、Luminex多因子检测、抗体芯片、PCR Array、ELISA、Elispot、PLA蛋白互作、多色免疫组化、DSP空间多组学等30多个,建立起了一套涵盖基因、蛋白、细胞以及组织水平实验的完整检测体系。
原文点击:Nature 重磅研究 | Naveni®PLA 原位技术揭秘 RAD52 维持复制叉稳态的分子新机制