

Cre-Lox系统新玩法:DiCre、Split-Cre与FISC三种工具小鼠精准调控方案
2026-07-01 来源:南模生物 点击次数:22你一定听说过Cre-Lox重组系统,无论你是否直接进行过基因操作。由于Cre-Lox系统具有操作简单、重组率高的优点,如今已经成为体内外遗传操作的强有力工具。利用Cre-Lox系统,可以在特定细胞、组织或整个生物体,甚至在特定时间点敲除或表达某个基因,实现对特定基因的时空特异性操作,这对基因功能的研究和人类疾病动物模型的建立都具有深刻影响。
传统的Cre-Lox系统虽然可以特异性地标记目的细胞的靶基因,同时后续也可以通过CreERT2的方式实现时间上对Cre的控制。但是在使用过程中还是常常遇到各种问题:单基因是否能特异性地标记某类细胞?他莫昔芬诱导要不要考虑代谢问题、毒性问题呢?万一这类细胞找不到合适的靶基因又该怎么办呢?
今天小编为您细说Cre-Lox系统的老套路和新玩法。
老套路
· Cre重组酶
环化重组酶(Cre, cyclization recombinase),是酪氨酸位点特异性重组酶之一,能催化两个DNA识别位点之间的位点特异性重组。Cre重组酶来源于P1噬菌体,由343个氨基酸组成,能特异性地识别loxP位点。
· loxP位点
Cre重组酶识别的回文DNA位点,也叫loxP(locus of X-over P1)位点,长34 bp,其特征结构为ATAACTTCGTATA-NNNTANNN-TATACGAAGTTAT,两边反向互补的13个碱基为Cre重组酶的识别序列。
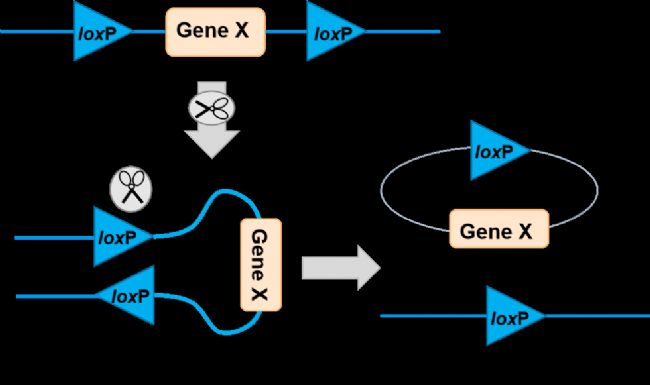
图1. Cre重组酶的识别序列
· 基本应用
合理利用Cre-Lox系统可以实现对基因的操控,根据您的需求进行基因的条件性表达或条件性敲除。关于Cre-Lox系统的原理与基本应用,我们官网上有详细介绍哦,链接放在阅读原文啦,感兴趣的点击下方了解吧。
新玩法
基于小鼠的Cre-Lox系统应用
1. DiCre系统——雷帕霉素诱导的时间调控
早在2003年Jean-Paul Herman团队就提出,Cre酶由于缺乏对其活动的有效时间控制,它的实用性仍然受到限制。后来他们开发了DiCre,一种用于Cre的可调节片段互补系统。
· 设计原理
Cre酶被分成两个部分,分别与FKBP12(FK506结合蛋白12)和FRB(FKBP12-雷帕霉素相关蛋白的结合域)融合。
· 诱导机制
通过雷帕霉素的诱导快速重组,利用雷帕霉素与FKBP12和FRB结构域结合后,可以直接诱导这两个结合结构域的异二聚化,实现Cre酶的时间调控。
(Nicolas Jullien et al., Nucleic Acids Res. 2003)
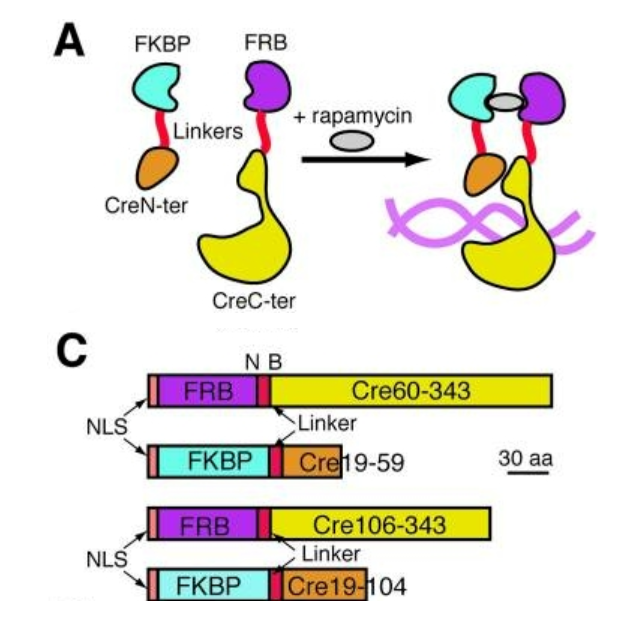
图2. DiCre系统原理示意图
2009年Frank Kirchhoff团队针对Cre/loxP系统单基因的表达模式存在不特异性的问题,且单启动子不足以从遗传学上定义不同的神经细胞类型,他们在上述研究基础上改进了DiCre,并重新定义为"Split-Cre"系统。
· 结构设计
具有N端或C端Cre片段的分别融合到酵母转录激活因子GCN4的组成型二聚化卷曲螺旋亮氨酸拉链域中(两个拉链区可以由α螺旋一侧的Leu之间的疏水作用形成拉链,并以α螺旋超卷曲的连接方式形成二聚体),分别定义为NCre和CCre。
· 工作原理
通过GCN4的二聚化实现Cre的重组。在此结构的基础上连接上不同基因的启动子,实现只有在发育过程中NCre和CCre同时表达的细胞才具有Cre活性。
(Johannes Hirrlinger et al., PLoS One. 2009)
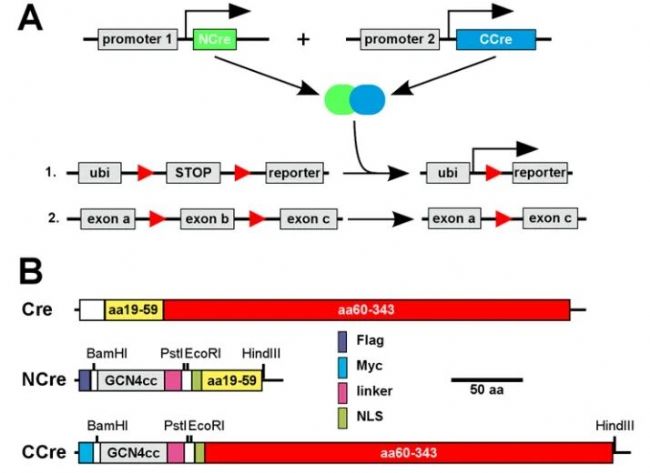
图3. Split-Cre互补原理示意图
· 技术对比
两个研发团队通过不同的方式都实现了对Cre活性的调控,"Split-Cre"系统可以说是在DiCre基础上做了完善,实现了Cre作用的更精准更特异,但是却舍弃了DiCre实现对Cre时间上的调控的设计初衷。随着技术的发展CreERT2进入了人们的视野,但是化学诱导不可避免存在一定的毒性,这个问题对一些药物敏感性实验成为了致命打击。
3. FISC系统——远红光诱导的无毒性调控
2020年Haifeng Ye团队针对化学诱导毒性问题,并结合前人研究,提出了新的系统:远红光诱导分裂Cre-loxP系统(FISC系统)。
· 设计思路
该系统将Cre的N端融合到Coh2结构域,Cre的C端与DocS结构域融合,并由远红外光(FRL)响应启动子pFRLx驱动。
· 激活机制
在FRL光照下,基于Coh2和DocS结构域的强亲和力,当将两个片段结合在一起时,Cre酶才能拥有重组酶活性,行使相应功能。
(Jiali Wu et al., Nat Commun. 2020)
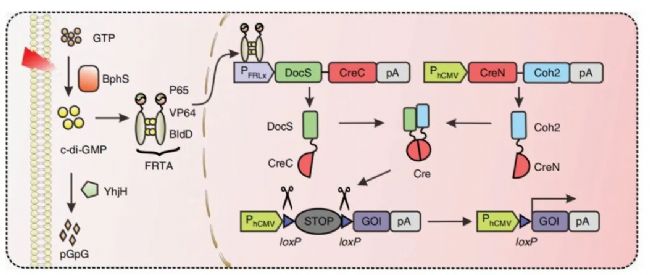
图4. FISC系统设计与优化示意图
以上研究都是基于转基因小鼠为研究模型对Cre-loxP系统的不同应用。南模生物可提供超过300多种自主产权Cre工具小鼠,覆盖全身各类型的细胞组织,满足科研人员多样化的需求。同时,南模生物长期提供Cre工具小鼠定制服务,可按具体实验要求进行定制,满足不同实验室需求。
南模生物项目交付流程
基于AAV的Cre-Lox系统应用
固然,根据具体实验方案定制培育Cre小鼠是更贴合实验需求的方式,但定制Cre小鼠不仅费时而且昂贵,且管理多种Cre小鼠品系更是繁琐易错,那么这一问题又该怎么解决呢?
· AAV载体的优势
重组腺相关病毒载体(AAV)介导的Cre-loxP系统很好地解决了上述问题。将目的元件包括启动子和增强子,以及内含子、微小RNA识别序列和内部核糖体进入位点(IRES或2A),重组酶(Cre、CreER、Flp或FlpER),翻译后调控元件,如WPRE或3'-UTR等,构建出特异性组织或者细胞表达的Cre工具小鼠。
(Leila Haery et al., Front Neuroanat. 2019)
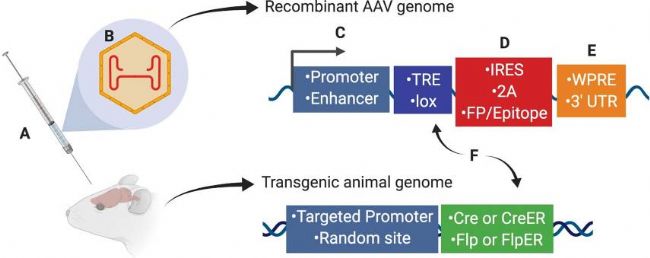
图5. AAV介导的神经元靶向与操作实验设计
· AAV系统的局限性
但是AAV转导的效率和特异性也会受到很多因素的影响,例如给药的途径、AAV血清型、AAV滴度和给药剂量等。同时哺乳动物的基因转录的调控元件通常分布在数千到数十万个碱基上,但是AAV基因组的包装能力是有限的(小于5 kb),所以识别与AAV兼容的启动子一直具有挑战性。这些也都使AAV介导Cre-loxP系统受到了一定的限制,所以到底是用小鼠或是AAV还是要视情况而定的。
南模生物的验证数据
南模生物利用AAV介导的Cre-Flox系统构建了丰富的基因敲除小鼠模型,部分验证数据如下:
小鼠肺癌模型
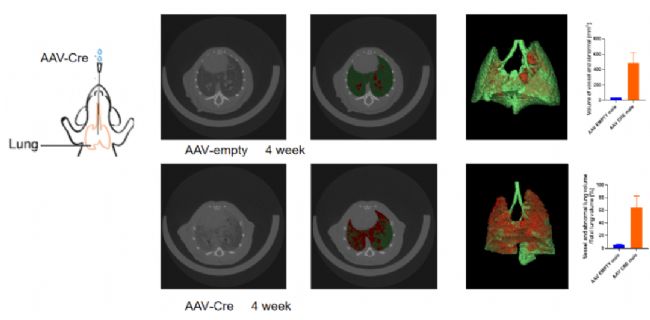 图6. 由气管雾化注射AAV-Cre病毒至KL小鼠(NM-CKO-234719)肺部,使Cre在小鼠肺部特异性表达,诱导Stk11表达缺失与Kras-G12D突变,可诱发肺癌发生。
图6. 由气管雾化注射AAV-Cre病毒至KL小鼠(NM-CKO-234719)肺部,使Cre在小鼠肺部特异性表达,诱导Stk11表达缺失与Kras-G12D突变,可诱发肺癌发生。小鼠PKD运动障碍模型
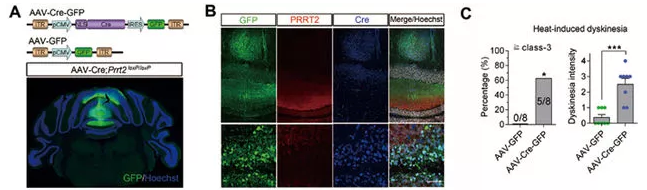 图7. 腺相关病毒介导Cre在小脑内局部敲除Prrt2表达,导致热诱导性运动障碍的发作。
图7. 腺相关病毒介导Cre在小脑内局部敲除Prrt2表达,导致热诱导性运动障碍的发作。南模生物Find Cre® 数据库
南模生物深耕基因修饰动物模型行业二十余年,为快速精准地选择合适Cre工具小鼠,特推出Find Cre®数据库。该数据库包含超过400多种自主产权Cre/Dre重组酶工具小鼠,以小鼠组织、器官和系统为主线,可分为肺、肝、胃肠道、乳腺、胰腺、感觉器官、心血管系统、免疫系统、神经系统、泌尿生殖系统、骨骼肌系统以及其他组织器官。大家只要选择自己感兴趣的组织器官,就可以查看该组织下可用的工具小鼠。
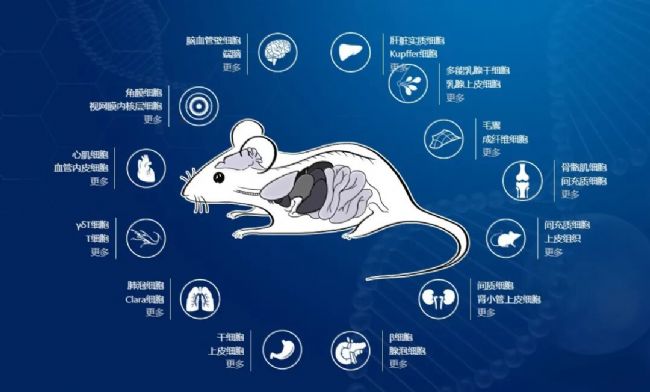
图8. 南模生物Find Cre®数据库
参考文献
Nicolas Jullien et al. Regulation of Cre recombinase by ligand-induced complementation of inactive fragments. Nucleic Acids Res. 2003 Nov 1;31(21):e131. doi: 10.1093/nar/gng131.
Johannes Hirrlinger et al. Split-cre complementation indicates coincident activity of different genes in vivo. PLoS One. 2009;4(1):e4286. doi: 10.1371/journal.pone.0004286. Epub 2009 Jan 27.
Jiali Wu et al. A non-invasive far-red light-induced split-Cre recombinase system for controllable genome engineering in mice. Nat Commun. 2020 Jul 24;11(1):3708. doi: 10.1038/s41467-020-17530-9.
Leila Haery et al. Adeno-Associated Virus Technologies and Methods for Targeted Neuronal Manipulation. Front Neuroanat. 2019 Nov 26:13:93. doi: 10.3389/fnana.2019.00093. eCollection 2019.
Tan GH, Liu YY, Wang L, et al. PRRT2 deficiency induces paroxysmal kinesigenic dyskinesia by regulating synaptic transmission in cerebellum. Cell Res. 2018;28(1):90-110. doi:10.1038/cr.2017.128
关于我们
上海南方模式生物科技股份有限公司(Shanghai Model Organisms Center, Inc.,简称"南模生物"),成立于2000年9月,是一家上交所科创板上市高科技生物公司(股票代码:688265),始终以编辑基因、解码生命为己任,专注于模式生物领域,打造了以基因修饰动物模型研发为核心,涵盖多物种模型构建、饲养繁育、表型分析、药物临床前评价等多个技术平台,致力于为全球高校、科研院所、制药企业等客户提供全方位、一体化的基因修饰动物模型产品解决方案。