

CD3/CD19双特异性抗体研发进展与人源化小鼠模型选择指南
2026-07-15 来源:南模生物 点击次数:422025年10月1日,阿斯利康注册了CD3/CD19双抗AZD0486(Surovatamig)治疗类风湿性关节炎或系统性红斑狼疮的一期临床试验。该试验计划入组48例类风湿性关节炎或系统性红斑狼疮患者,预计2028年完成。
AZD0486由Tenebio研发,阿斯利康在2022年7月以12.65亿美元收购。与其相似的情况屡屡发生:默沙东在2024年8月耗费13亿美元收购CD3×CD19双抗CN201;罗氏披露了在研自免双抗管线CD3/CD19双抗RG6382。
截至2026年,该赛道继续加速:德琪医药ATG-201(CD19×CD3 TCE)已在中国获批IND,适应症为自身免疫疾病;Xencor的XmAb657(B细胞清除型TCE)已于2025年底进入临床阶段;Abogen在AACR 2026报告了ABO2203(mRNA编码的CD3×CD19 TCE)的首次人体研究初步结果。
为何CD3/CD19靶点组合能够成为领域内关注的焦点?下面我们逐一拆解。
CD3概述
CD3是表达于T细胞膜上的关键表面标志物,在T细胞活化中扮演核心角色,并因其功能的不可或缺性而成为极具潜力的药物靶点。
结构组成:
CD3由四条蛋白质链(CD3γ、CD3δ、CD3ε和CD3ζ)构成复合体。其中CD3γ、CD3δ和CD3ε的胞外结构域两两配对,形成CD3γε与CD3δε两对异二聚体;它们与T细胞受体(TCR)以及同源二聚体CD3ζζ共同组装成完整的TCR/CD3复合物。
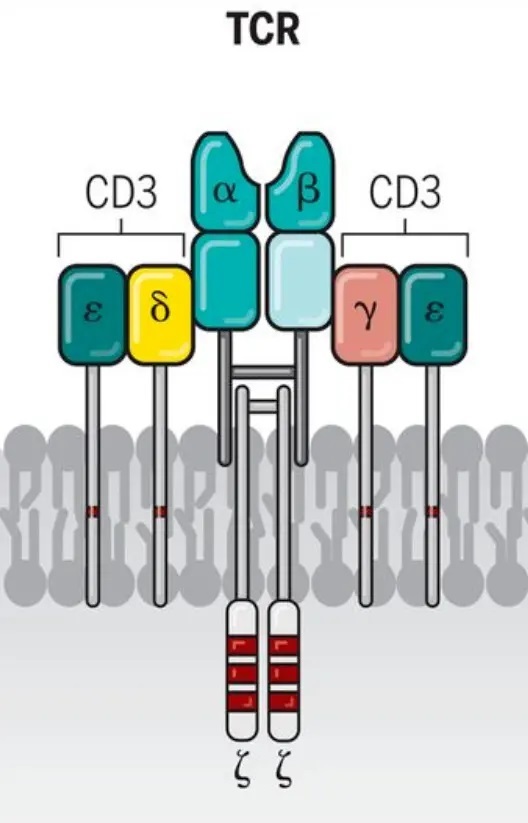
功能要点:
· 信号转导——CD3胞内区含有免疫受体酪氨酸激活基序(ITAM),尤其在CD3ζ链上分布有多个。当TCR识别抗原后,CD3负责启动下游信号转导,直接调控T细胞的活化、增殖与分化。
· 发育调控——该复合物在T细胞的发育成熟过程中也起着至关重要的调控作用。
CD19是一种分子量约为95 kDa的跨膜糖蛋白,属于免疫球蛋白超家族成员。作为B细胞谱系中至关重要的表面标志物,它在从早期前B细胞到成熟B细胞的发育全程中稳定表达,但其表达会随着B细胞终末分化为浆细胞而消失。
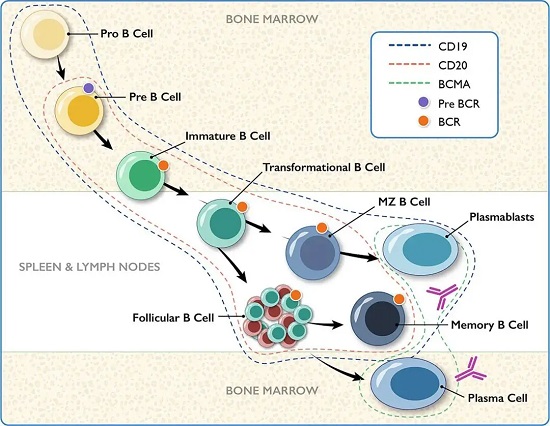
在肿瘤治疗中的意义:
CD19在绝大多数B细胞恶性肿瘤中呈现高表达,尤其是在前体B细胞急性淋巴细胞白血病(BCP-ALL)中。此外,它也广泛高表达于ALL、CLL及多种B细胞淋巴瘤,为靶向治疗提供了良好的窗口。
在自身免疫疾病中的角色:
CD19作为B细胞信号转导的关键共受体,其表达与功能的异常在系统性红斑狼疮(SLE)等多种自身免疫性疾病的发病机制中同样扮演重要角色。以SLE为例,CD19与BCR(B细胞受体)及其他表面分子(如CD21)形成复合物,共同构成一个关键的信号放大模块。在SLE患者中,B细胞表面的CD19表达水平往往上调或其信号通路过度活跃,导致B细胞对抗原刺激的应答阈值降低,即使面对微弱信号也能产生过度活化,进而导致病理性自身抗体(如抗dsDNA抗体、抗Sm抗体等)的大量产生。这些自身抗体是驱动SLE免疫复合物形成、沉积并引发多组织器官(如肾脏、皮肤、关节)炎症损伤的直接效应因子。
结合二者的TCE双抗
双特异性抗体由于含有两种抗体的特异性,可以结合同一种抗原的两种表位或两种不同抗原的表位。效应位点起到连接募集效应细胞、药物分子或病毒的作用;靶点位点可以靶向到分子或细胞等。
T细胞衔接器(TCE)双抗指的是一种能同时结合肿瘤相关抗原(TAA)和T细胞上CD3的双特异性抗体。其设计目的是将T细胞定向到靶细胞(肿瘤细胞或致病B细胞),从而使T细胞能有效地杀伤目标细胞。
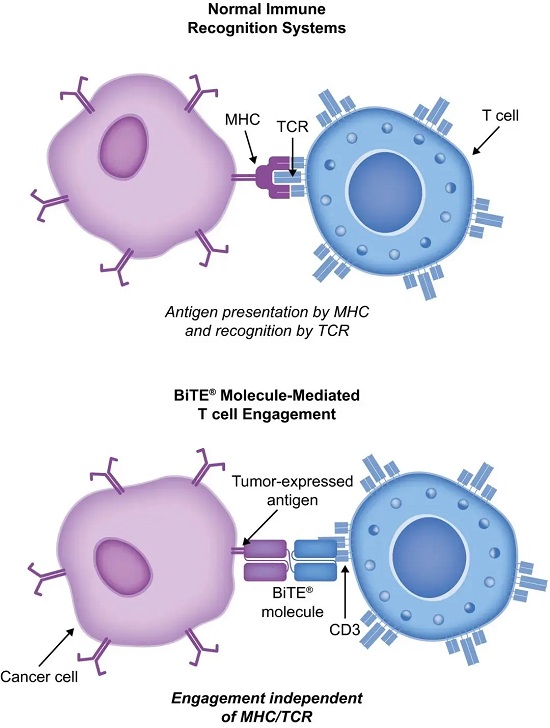
由于CD19在B细胞恶性肿瘤与B细胞相关自免疾病中均起到关键作用,CD3/CD19双抗的适应症也不断扩大。已获批用于治疗耐药急性淋巴细胞白血病的贝林妥欧单抗(Blinatumomab),也在研究阶段被证实可用于治疗难治性类风湿性关节炎、系统性红斑狼疮等自身免疫疾病。针对自身免疫疾病模型的临床前CRO评价也成为该类药物研发的重要环节。
南模生物相关小鼠模型
基于CD3/CD19双特异性抗体在肿瘤及自身免疫性疾病治疗领域的广阔前景,其临床前研究的关键环节之一在于选用能准确模拟人体免疫反应及疾病进展的动物模型。南模生物可为您提供C57BL/6背景与BALB/c背景的hCD3EDG/hCD19双靶点人源化小鼠模型,为您的抗体筛选、药效评估及作用机制研究提供可靠且高效的工具支持。
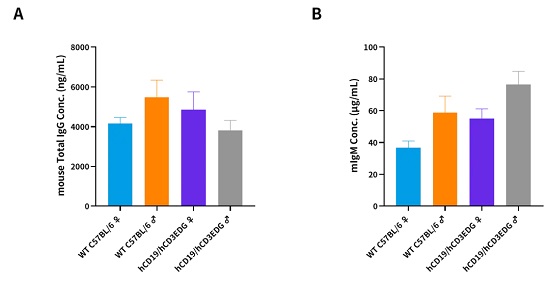
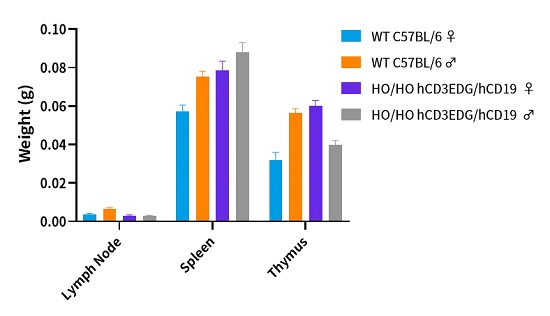
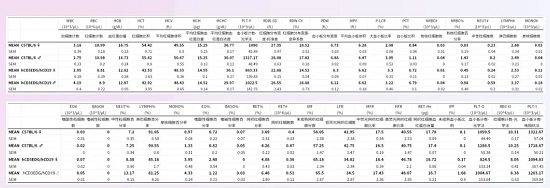
hCD3EDG/hCD19(C57BL/6)小鼠免疫表型正常
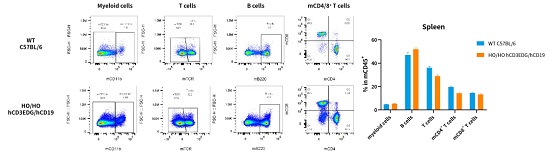
hCD3EDG/hCD19(BALB/c)小鼠免疫表型正常

南模生物相关药效评价服务
在hCD3EDG/hCD19双靶点人源化小鼠的基础上,南模生物可为您提供疾病造模与药效评价等多种临床前CRO服务,为您的相关药物开发提供支持。
hCD3EDG/hCD19(BALB/c)可响应CD19/CD3双抗药物
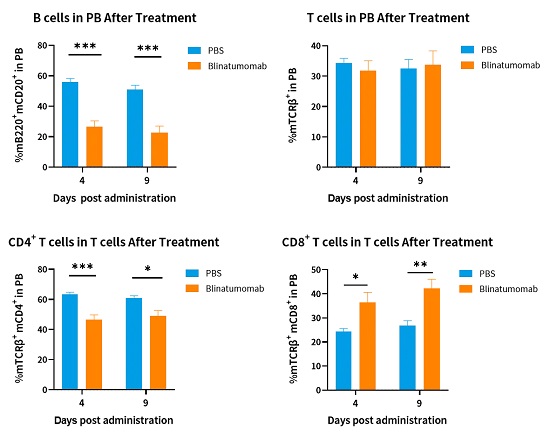
:B细胞数量明显减少,T细胞总数未改变,但CD8+ T细胞比例增加。
hCD3EDG/hCD19(BALB/c)小鼠可构建SLE自身免疫疾病模型,并用于SLE药效检测
造模方法:
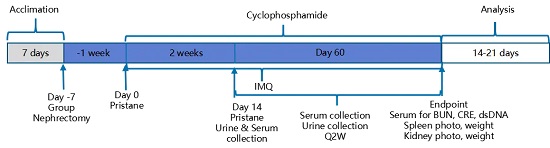
分组情况:
· G1对照组: 未处理的hCD3EDG/hCD19(BALB/c)小鼠
· G2疾病组: 使用IMQ+Pristane诱导发病的肾切除hCD3EDG/hCD19小鼠
· G3给药组: 在疾病小鼠基础上给药cyclophosphamide
药效数据:
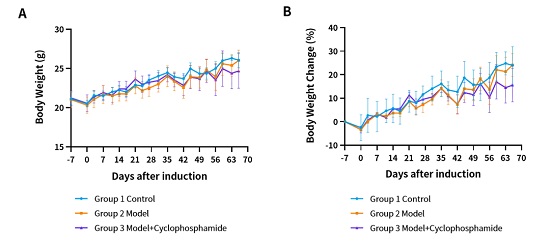
(A) 体重。(B) 体重变化。
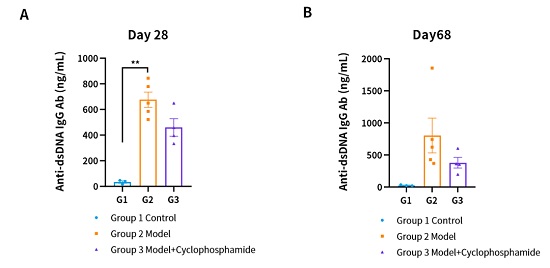
,而在给药后有所回落。(A) 第28天血清抗dsDNA IgG Ab。(B) 第68天的血清抗dsDNA IgG Ab。
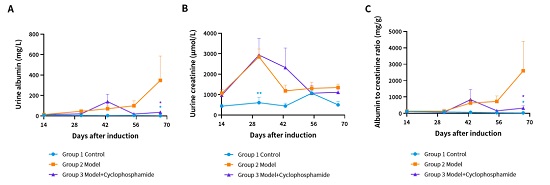
,而在给药后回落至对照组水平。(A) 尿白蛋白。(B) 尿肌酐。(C) 白蛋白与肌酐比值。(* P < 0.05, ** P < 0.01 Vs Group2)。
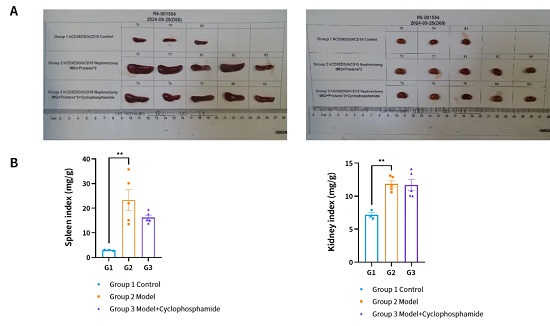
,在给药后脾脏大小有所回落,但肾脏大小未出现明显差异。(A) 脾脏照片和肾脏照片。(B) 脾指数和肾指数。(** P < 0.01 Vs Group2)。
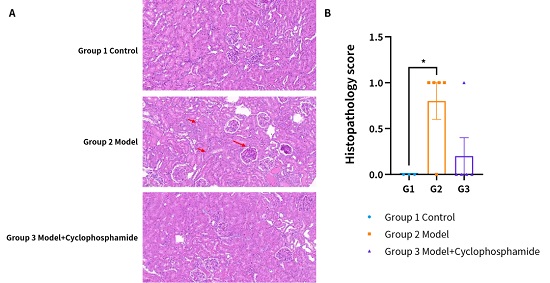
:疾病造模后系膜细胞、内皮细胞增生,伴多形核中性粒细胞浸润,常伴有白金耳或新月体形成,出现明显的狼疮性肾炎表型;而在给药后该表型出现明显缓解。(A) 代表性H&E图像。(B) 组织病理学评分。(* P < 0.05 Vs Group2)。
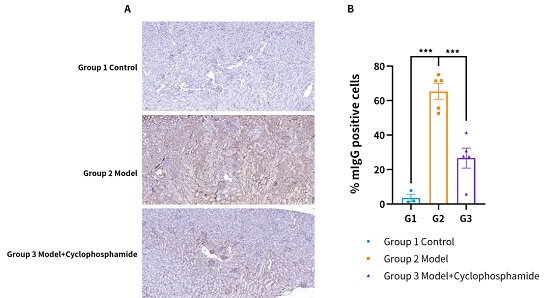
:疾病造模后肾脏IgG呈强阳性表现;而在给药后IgG阳性信号减少。(A) 系统性红斑狼疮模型的代表性IHC图像。(B) mIgG阳性细胞百分比。
Reference
Carl H. June et al., CAR T cell immunotherapy for human cancer. Science 359, 1361-1365(2018). DOI:10.1126/science.aar6711
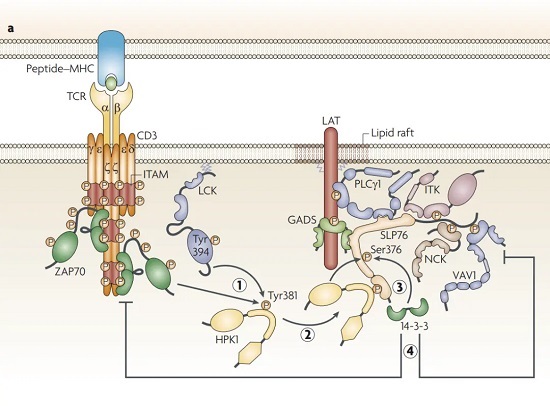
Acuto, O., Bartolo, V. & Michel, F. Tailoring T-cell receptor signals by proximal negative feedback mechanisms. Nat Rev Immunol 8, 699–712 (2008).
Williams JP, Meyers JA. Immune-mediated inflammatory disorders (I.M.I.D.s): the economic and clinical costs. Am J Manag Care. 2002 Dec;8(21 Suppl):S664-81.
Einsele H, et al. The BiTE (bispecific T-cell engager) platform: Development and future potential of a targeted immuno-oncology therapy across tumor types. Cancer. 2020 Jul 15;126(14):3192-3201.
关于我们
上海南方模式生物科技股份有限公司(Shanghai Model Organisms Center, Inc.,简称"南模生物"),成立于2000年9月,是一家上交所科创板上市高科技生物公司(股票代码:688265),始终以编辑基因、解码生命为己任,专注于模式生物领域,打造了以基因修饰动物模型研发为核心,涵盖多物种模型构建、饲养繁育、表型分析、药物临床前评价等多个技术平台,致力于为全球高校、科研院所、制药企业等客户提供全方位、一体化的基因修饰动物模型产品解决方案。
欢迎通过以下方式联系我们:
微信公众号「南模生物SMOC」在线咨询
拨打400-728-0660
点击文末「阅读原文」