

文献解读:脑内色胺信号轴的发现揭示睡眠压力调控通路
2026-06-26 来源:本站 点击次数:63
2026年6月19日,中国科学院脑科学与智能技术卓越创新中心张哲研究员团队牵头,联合穆宇研究员团队、复旦大学袁鹏团队、陆军军医大学胡志安团队、上海科技大学刘志杰与华甜课题组、复旦大学华山医院郁金泰临床团队等多家单位协同在《Nature Neuroscience》期刊发表了题为:“Tryptamine from wake-active monoaminergic neurons regulates sleep homeostasis”的研究论文。汇集神经生物学、结构生物学、临床遗传学多学科研究力量开展跨物种系统验证。研究团队依托靶向代谢组、基因编码荧光探针、单细胞转录组、CRISPR 基因编辑、光遗传与膜片钳等多维度实验技术,以夜行小鼠、昼行巴马小型猪为动物模型层层递进开展机制解析;首次证实色胺(TrpA)是由清醒型单胺能神经元活性依赖合成释放的内源睡眠信使,脑脊液色胺浓度可精准、独立于昼夜节律与应激水平表征睡眠压力;进一步鉴定孤儿受体 GPR139 为色胺特异性内源受体,明确下丘脑视前区表达 GPR139 的 GABA 抑制神经元是机体感知睡眠压力的核心效应细胞,完整阐明 “蓝斑 / 中缝背核单胺能神经元分泌色胺 — 脑脊液扩散 — 激活 POA-GPR139 神经元 — 抑制全脑觉醒环路、驱动深度非快速眼动睡眠” 完整调控通路;同时证实 TrpA-GPR139 信号是睡眠剥夺后代偿性睡眠反弹的必需通路,可透过血脑屏障的 GPR139 小分子激动剂能够提升睡眠时长与深度,人类 AADC、GPR139 基因多态性也与失眠发病风险存在关联。该研究补齐睡眠稳态理论缺失的关键分子链条,同时为稳态受损型失眠开辟区别于传统广谱镇静药物的全新靶向治疗思路,为睡眠障碍病理机制解析与创新助眠药物研发提供坚实基础理论与实验依据。
01、研究方法
1.1 模式动物与造模体系
实验同时采用夜行 C57BL/6 小鼠、昼行巴马小型猪两大哺乳动物模型,消除昼夜节律作息差异带来的实验偏差;构建多种基因工具小鼠(Th-Cre、Sert-Cre、DAT-Cre、Gpr139-Cre)实现特定神经元特异性标记;通过脑室微量注射、核团局部给药、全身药物干预三种给药方式区分全身与局部作用;构建睡眠剥夺、自由睡眠、恢复睡眠三组标准睡眠造模体系;借助 CRISPR-Cas9 病毒实现下丘脑视前区(POA)局部、全脑两种 GPR139 基因敲除模型。
1.2 代谢组与分子互作检测手段
采用液相色谱 - 串联质谱完成脑脊液靶向代谢组初筛,超高效液相色谱定量目标分子浓度;体外构建 GPR139 过表达细胞系开展钙流功能实验;依托蛋白晶体结构开展分子对接,直观验证色胺与 GPR139 的直接结合作用;使用 AADC 酶抑制剂、色胺原液、GPR139 选择性激动剂完成药理功能验证。
1.3 活体动态观测技术
自主构建比例型色胺荧光遗传探针,搭配光纤光度成像实时观测活体神经元色胺释放动态;使用 GCaMP6s 钙探针记录睡眠调控神经元的活体放电活动;多导睡眠记录仪同步采集脑电波、肌电信号,自动划分清醒、NREM、REM 三种脑状态;离体脑片膜片钳记录神经元静息电位、放电阈值,量化细胞兴奋性变化。
1.4 神经元操控与单细胞测序
运用光遗传、化学遗传双向激活 / 抑制单胺能神经元、POA 睡眠神经元;借助破伤风毒素阻断神经元突触释放,验证分子释放依赖囊泡转运;对睡眠剥夺、恢复期小鼠 POA 脑区开展单细胞 RNA 测序,筛选响应睡眠压力的特异性神经元亚群;免疫荧光、RNA 原位杂交完成蛋白、基因的脑内定位染色。
02、研究内容
2.1 色胺是脑脊液中表征睡眠压力的内源促眠分子
研究首先采集自由睡眠、睡眠剥夺两组小鼠脑脊液开展代谢组筛选,在众多神经代谢物中发现色胺(TrpA)含量存在显著差异。整体围绕色胺与睡眠压力的相关性设计多组验证:完成小鼠脑脊液代谢差异初筛,追踪完整光暗周期内脑脊液色胺浓度变化,结果显示动物清醒阶段色胺持续累积,进入睡眠周期后含量逐步回落;设置短时睡眠剥夺干预,人为提升睡眠压力后,脑脊液色胺同步出现明显上升。为排除光照、内源昼夜节律干扰,采用全黑暗无光照饲养小鼠,仅依靠滚轮活动区分清醒、休息时段,色胺浓度依旧仅和动物清醒活动量正相关;同时检测睡眠剥夺小鼠血清皮质醇水平,证明色胺升高并非应激反应导致。 为验证物种保守性,引入昼行巴马小型猪重复时序检测,和人类相近的作息下,色胺依旧随清醒时长升高、入睡后下降,证明该规律在哺乳动物中通用。后续开展脑室注射功能验证:向小鼠脑室内梯度输注色胺后,动物 NREM 睡眠时长、单次睡眠片段长度、脑慢波 δ 功率同步上升,呈现剂量依赖性促眠效果;无论在小鼠固有清醒暗期、睡眠光期给药,色胺均能有效提升睡眠总量;不同基线睡眠压力下给药结果显示,色胺的促眠效应与机体本身的睡眠压力存在协同作用,直接证明脑脊液色胺具备内源促眠活性,可精准反映睡眠压力高低。
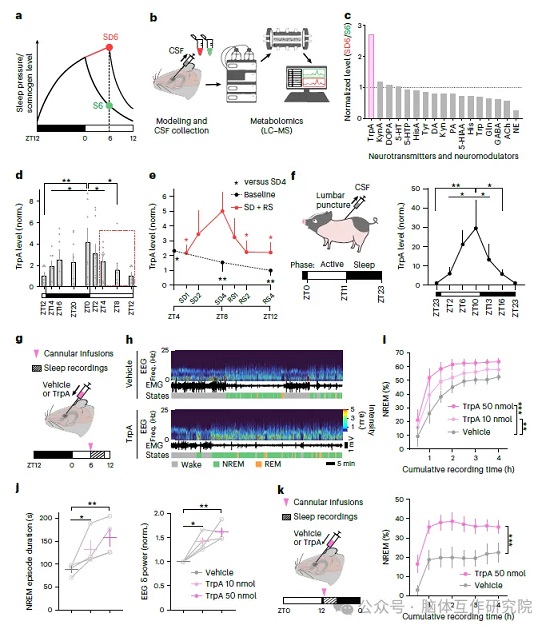
2.2 清醒状态下单胺能神经元活性依赖性合成分泌色胺
明确色胺功能后,实验追溯色胺的脑内合成来源。 阐明合成通路:色氨酸经 AADC 酶脱羧生成色胺,免疫荧光染色显示 AADC 酶高度富集于蓝斑 LC、中缝背核 DR、腹侧被盖区 VTA 三类单胺能神经元;全身注射 AADC 抑制剂后,睡眠剥夺无法提升脑脊液色胺,睡眠代偿性反弹同步消失,证明 AADC 酶是色胺合成必需条件。 为直观观测活体神经元色胺释放动态,团队改造得到高特异性色胺荧光探针,该探针仅响应色胺,对色氨酸、5 - 羟色胺等相似代谢物无明显荧光响应,具备毫秒级动态检测能力。将探针通过病毒特异性表达在三类单胺能神经元内,光纤光度成像结果显示:小鼠清醒阶段,LC、DR、VTA 神经元轴突持续出现探针荧光快速下降信号,代表色胺大量释放;进入 NREM、REM 睡眠后,释放信号大幅减弱;沉默突变对照探针无对应动态变化,排除 pH、运动等无关干扰。通过光遗传分别激活 LC、DR 神经元,激活后脑脊液色胺含量显著上升;向单胺能神经元表达破伤风毒素阻断突触囊泡释放,清醒状态下的色胺释放信号完全消失,证明色胺依靠神经元活动、突触释放进入脑脊液。同时实验观测丘脑、结节乳头核等其他清醒核团,其神经元不会产生色胺释放信号,确定 LC、DR 为色胺主要分泌细胞,仅清醒活跃的单胺能神经元可活动依赖性生成色胺。
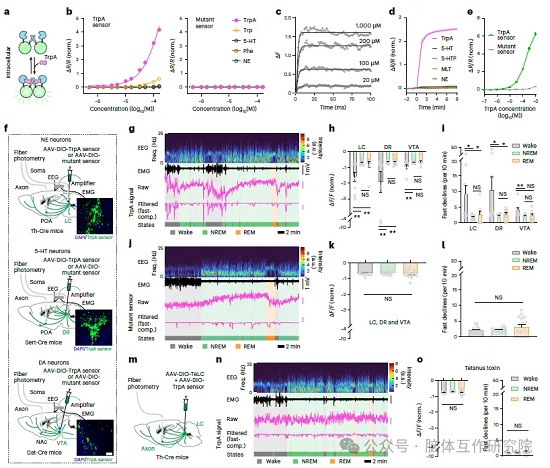
图2:清醒状态下单胺能神经元释放色胺
2.3 色胺通过下丘脑 POA 区 GPR139 受体调控睡眠神经元兴奋性
本部分锁定色胺在大脑的下游作用靶点。首先向小鼠脑室输注色胺后检测全脑 c-Fos 激活标记,下丘脑视前区 POA 神经元激活程度显著高于其他脑区,确定 POA 为色胺核心作用区域。对睡眠剥夺、恢复期小鼠 POA 脑组织开展单细胞 RNA 测序,对比两组细胞基因表达差异后发现:一类 GABA 抑制神经元(i2 亚群)在高睡眠压力恢复期大量表达即刻早期激活基因,该细胞高表达孤儿受体 GPR139,是唯一同步响应睡眠压力、高表达 GPR139 的神经元亚型。 体外功能实验验证分子互作:提取睡眠剥夺小鼠脑脊液处理过表达 GPR139 的细胞,可诱发明显钙信号;分子对接模拟直观显示色胺能够稳定结合 GPR139 正构结合口袋,梯度色胺原液处理细胞可剂量依赖性激活受体钙信号,证实 GPR139 是色胺内源受体。进一步解析胞内信号通路:GPR139 激活下游 Gq/11 蛋白,抑制 GIRK 钾通道,降低神经元放电阈值。向小鼠 POA 局部共注射色胺与 Gq 通路抑制剂、GIRK 通道激活剂,色胺原本的促眠效果完全消失,完整阐明色胺 - GPR139 调控神经元兴奋性的分子通路。使用 Gpr139-Cre 工具鼠完成细胞定位染色,GPR139 阳性神经元集中分布于腹外侧视前区 VLPO,恢复期该类神经元 c-Fos 阳性比例大幅上升,印证其睡眠压力感知功能。
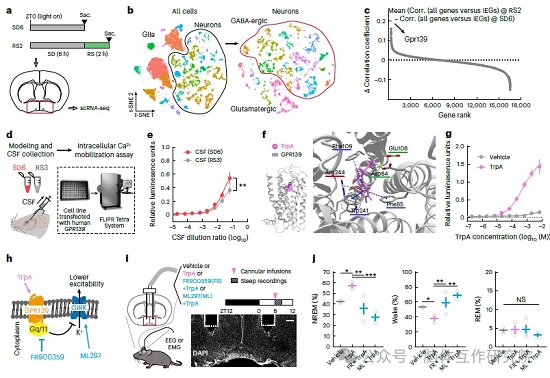
图3:鉴定下丘脑视前区 GPR139 为色胺的特异性受体
2.4 POA 区 GPR139 神经元是睡眠压力核心效应单元,通路缺失直接破坏睡眠稳态
围绕 GPR139 神经元的兴奋性与睡眠压力关联开展验证:分离自由睡眠、睡眠剥夺小鼠 POA 脑片进行膜片钳记录,睡眠剥夺组 GPR139 神经元动作电位发放阈值更低、同等刺激下放电频率更高;脑片外源性施加色胺,同样会降低细胞放电阈值,证明高色胺、高睡眠压力会提升该类神经元兴奋性。活体钙成像实验同步记录神经元钙信号与睡眠状态:清醒向 NREM 睡眠切换时,GPR139 神经元钙信号显著上升;NREM 向清醒切换时钙信号快速回落;睡眠压力更高的实验早期,神经元睡眠特异性激活幅度更强,随睡眠推进、压力消散,激活幅度逐步减弱。光遗传实验双向操控细胞:激活 GPR139 神经元直接延长 NREM 睡眠、缩短清醒时长;抑制该神经元会快速唤醒小鼠、瓦解持续睡眠。神经示踪显示,POA GPR139 神经元广泛投射 LC、DR、VTA 等全脑觉醒核团,通过抑制觉醒环路稳定睡眠。
开展基因缺失功能验证:借助 CRISPR 病毒局部敲除小鼠 POA GPR139 基因后,脑室输注色胺不再具备促眠效果;正常小鼠睡眠剥夺后会出现代偿性深度恢复睡眠,而 POA、全脑 GPR139 敲除小鼠无睡眠反弹,睡眠片段化、深度睡眠大幅减少,证明 TrpA-GPR139 信号通路是维持睡眠稳态不可或缺的核心环节。最后开展药物转化验证:两种可穿透血脑屏障的 GPR139 小分子激动剂全身给药,野生小鼠 NREM 睡眠时长、慢波深度明显提升;POA GPR139 敲除小鼠给药后无任何睡眠改善,证实靶向该受体可模拟内源色胺的促眠作用。
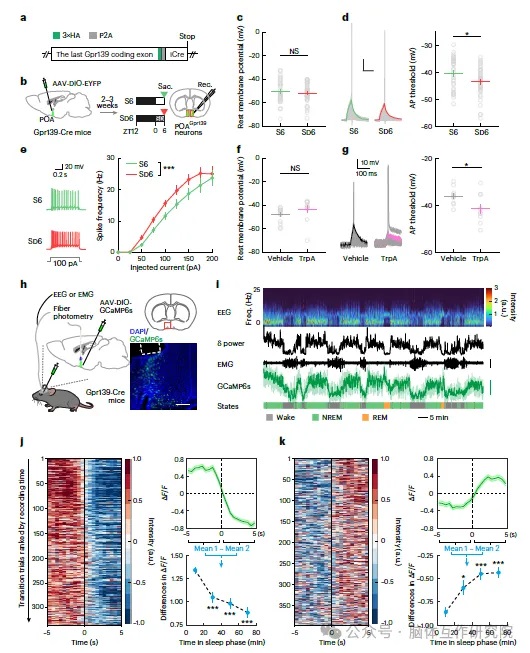
图4:色胺调控下丘脑视前区 GPR139 神经元兴奋性并编码睡眠压力
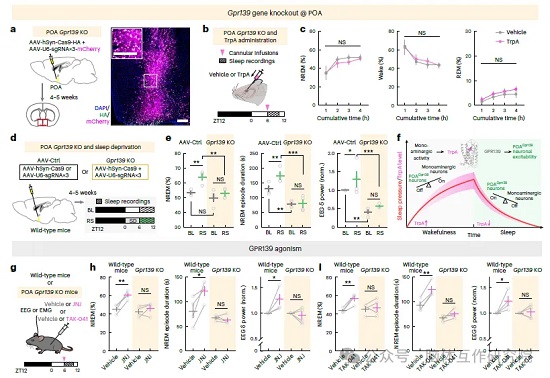
图5:GPR139 信号通路调控睡眠稳态
03、创新点
3.1 基础分子发现创新
首次发现色胺作为直接由觉醒型单胺能神经元合成的内源睡眠压力信使,解决持续近百年 “脑脊液促眠分子未知” 科学难题。区别于腺苷等不关联觉醒活动的传统促眠物质,色胺的生成严格绑定大脑清醒神经活动,完美对应 “越清醒、困意越强” 生理规律,完善经典双进程睡眠调控理论。
3.2 孤儿受体功能创新
首次明确孤儿受体 GPR139 的天然内源配体为色胺,阐明其在下丘脑睡眠中枢调控睡眠稳态的核心生理功能,打通 “单胺能神经元 — 色胺 —POA GPR139 神经元” 完整睡眠压力传导分子环路,填补孤儿受体生理功能研究空白。
3.3 检测工具技术创新
基于色氨酸传感蛋白定点突变,开发高选择性、高时间分辨率的色胺比例荧光探针,实现活体动物大脑神经元色胺释放动态可视化,为中枢单胺类中间代谢物的在体观测提供全新通用工具。
3.4 临床转化创新
提出 GPR139 作为失眠全新药物靶点,区别于传统广谱镇静药物,GPR139 激动剂依托人体天然睡眠压力调控通路起效,具备更贴合生理、低依赖的开发潜力;同时人类基因多态性数据佐证通路与睡眠疾病相关性,为失眠新药研发提供坚实基础研究支撑。
04、启发
4.1 科研选题启发
经典理论遗留的长期基础谜题具备极高研究价值,本研究从 “清醒为何累积困意” 这一经典生理疑问切入,从脑脊液代谢物中挖掘全新信号分子。针对已知通路的中间代谢产物开展功能挖掘,容易发现与主通路功能拮抗的全新生理效应,打破对神经递质仅关注终产物的固有研究思维。
4.2 实验设计与图表对应启发
文章实验与附图严格按照 “分子筛选→分子溯源→受体鉴定→细胞功能→通路缺失验证→药物转化” 递进顺序排布,每 1-2 张附图对应单一阶段实验结论,变量区分清晰、干扰对照完备。开展生理分子研究时,需分层设置昼夜节律、应激、细胞特异性对照,通过跨物种模型、离体 + 活体多维度实验相互佐证,保证结论特异性、可靠性。
4.3 多技术交叉应用启发
单一实验手段难以完整解析神经分子环路,本研究融合代谢组、基因编辑、活体光学成像、电生理、单细胞测序、分子模拟多类技术,各类实验结果相互印证,弥补单一技术局限。针对未知小分子,自主开发特异性荧光探针可实现活体动态观测,大幅提升机制解析深度。
4.4 转化医学研究启发
基础神经环路研究可直接落地临床药物开发,孤儿 G 蛋白偶联受体是精神、睡眠类疾病极具潜力的药物靶点。相较于无差别抑制全脑神经的传统药物,靶向人体天然内源调控通路的小分子激动剂,安全性、临床适配性更具优势,基础研究需同步兼顾基础机制与临床应用潜力。
4.5 科研客观素养启发
文章在讨论部分客观列明研究局限性,未夸大实验结论,如实指出外源色胺药效时长有限、人体临床样本数据缺失、GPR139 多效性待评估等不足,为后续细分方向预留研究空间。完整、辩证地解读实验结果,不片面放大单一正向结论,是基础科研必备严谨思维
参考文献:
Cao, H., Wang, K., Zhao, J. et al. Tryptamine from wake-active monoaminergic neurons regulates sleep homeostasis. Nat Neurosci (2026). https://doi.org/10.1038/s41593-026-02332-x
厚谱实验室(脑体互作研究—神经药效评价实验室)专注于脑体互作机制解析与神经药效精准评价的尖端实验技术高地。可以开展清醒动物小分子取样分析、神经递质代谢产物实时分析、癫痫睡眠脑电采集分析、在体电生理记录分析、脑机接口方案评价(大动物脑部手术)、神经调控、组织透明化、脊髓损伤康复验证实验电生理(SEP)、脑立体定位给药、自动无接触采血分析、葡萄糖钳夹实验、行为学迷宫实验和动物造模等各种动物实验服务!
-END-


想了解更多内容,获取相关咨询请联系
电 话:+86-0731-84428665
伍经理:+86-180 7516 6076
工程师:+86-180 7311 8029
邮 箱:consentcs@163.com
