

数字微流控无细胞系统的应用:高通量筛选植物球蛋白表达与稳定性条件
2026-07-16 来源:本站 点击次数:29摘要:借助数字微流控无细胞蛋白合成系统,高通量筛选明确植物球蛋白表达与稳定性的关键决定因子
关键词:无细胞蛋白合成、无细胞蛋白表达、数字微流控、植物球蛋白、高通量筛选、难表达蛋白、膜蛋白、蛋白稳定性、可溶性标签、半胱氨酸突变、血红素蛋白
一、植物球蛋白无细胞表达研究简述
1.1 本文摘要
血红素蛋白参与机体代谢与应激应答,但存在细胞毒性、折叠困难、传统活细胞表达产量低、通用方案无法跨物种复用,植物球蛋白(Pgbs)就是典型代表。本文利用数字微流控无细胞蛋白合成(DMF-CFPS)技术高通量并行筛选可溶性标签、氧化还原添加剂、半胱氨酸突变为丙氨酸三大变量,明确调控植物球蛋白无细胞蛋白表达与蛋白稳定性的关键决定因子。本研究选取两种优势突变体进行放大表达,成功获得可溶性脱辅基球蛋白。研究结果证实,无细胞蛋白合成(CFPS)(可点击查看)技术可快速、平行筛选蛋白理想表达条件,同时阐明保守半胱氨酸、氧化还原环境在植物球蛋白合成成熟过程中的作用。
1.2 研究对象
● 甜菜1型植物球蛋白野生型BvPgb 1.2及保守半胱氨酸突变体C86A;
● 全新鉴定8个燕麦植物球蛋白AsPgb,分3个同源组:1型两组(AsPgb1.1–1.3、AsPgb1.4–1.6)、3型一组(AsPgb3.1–3.2);3组定点突变体:AsPgb1.1-C70A、AsPgb1.5-C84A、AsPgb3.1-C161A(保守C→A替换);
二、传统蛋白表达体系局限与CFPS应用研究背景
2.1. 血红素/植物球蛋白表达瓶颈
植物球蛋白(Pgbs)参与NO清除、氧化应激调控,属于典型难表达血红素蛋白:脱辅基球蛋白易聚集、血红素对宿主细胞有毒、合成与血红素插入难以同步;针对一种Pgbs优化的表达条件无法直接套用至同源蛋白,传统体内表达需要数周试错,通量低。
2.2 保守半胱氨酸的未知作用
Pgbs序列高度保守半胱氨酸残基,易形成错配二硫键、引发氧化聚集;C突变为A可消除巯基,但该突变对不同Pgbs表达量、可溶性、热稳定性的影响缺乏系统对比。
2.3 传统筛选局限性
活细胞筛选受转化效率、细胞毒性限制;常规96孔CFPS体系试剂消耗大、条件并行数量少;数字微流控(电润湿操控纳升级液滴)实现全自动高通量并行筛选,48小时内可完成192组表达条件测试,大幅缩短优化周期。
三、实验方法(DMF-CFPS完整解决方案)
3.1 数字微流控卡盒工作原理
nuclera eProtein Discovery系统配套专用卡盒依托介电上电润湿实现纳升级液滴独立移动、混合、孵育、磁珠纯化;卡盒集成多试剂储液孔,可自动完成CFPS合成、BiFC荧光定量、链霉亲和素磁珠原位纯化整套流程;通过双分子荧光互补BiFC实时定量蛋白表达浓度。
3.2 eGene线性DNA模板构建
利用Nuclera eGene可溶性标签试剂盒PCR组装得到整套eGene线性表达模板。线性模板两端分别携带:
● N端:7种可溶性标签(P17/CUSF/FH8/TRX/SUMO/ZZ/SNUT)或无标签+3C蛋白酶酶切位点;
● C端:TEV酶切位点+GFP检测标签+Strep纯化标签;
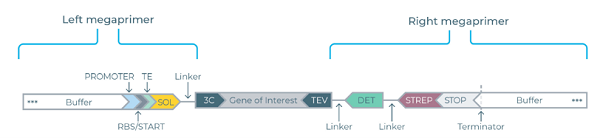
图1 eGene模板结构示意图
采用一步式PCR批量构建所有野生型与C→A突变eGene构建体,包含BvPgb 1.2、C86A、AsPgb 1.1、C70A、AsPgb 1.5、C84A、AsPgb 3.1、C161A。归一化浓度后进行卡盒上样。
3.3 配置无细胞反应液(CFB)
制备8组无细胞反应体系(CFB),每体系含无细胞核心试剂、定制氯化血红素补充液(用于辅因子插入测试)和不同功能的添加剂(标准缓冲液、PDI+GSSG、GSSG、TRXB1、DnaK分子伴侣混合液、金属辅因子混合液、Zn²⁺、3C蛋白酶),加样至卡盒对应孔。
3.4 两种卡盒筛选
● 3×8(甜菜BvPgb筛选):2种蛋白(WT/C86A)×(7种可溶性标签+无标签)×8组CFB,共128组条件;
● 6×4(燕麦AsPgb筛选):6种蛋白(3WT+3突变)×(3种可溶性小标签+无标签)×8组CFB,共192组条件;
启动可溶性蛋白筛选程序,自动筛选每组蛋白产量较高的组合(3×8选10组,6×4选5组)共30组,进行原位磁珠纯化,从而输出蛋白表达与纯化产量图谱。
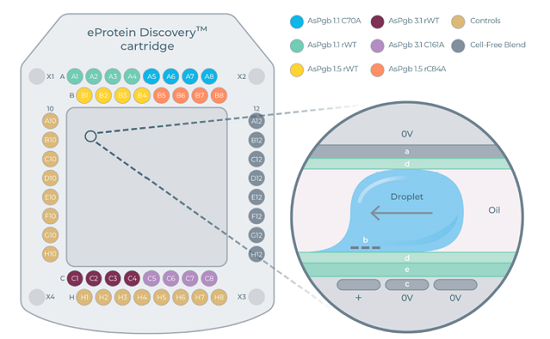
图2 用于无细胞蛋白合成(CFPS)的数字微流控卡盒
3.5 放大与蛋白表征
● 200 μL体系无细胞放大表达,Strep亲和纯化,SDS-PAGE验证;
● 质谱光度MP:测定蛋白寡聚状态;
● 纳米差示扫描荧光nanoDSF:测定蛋白熔解温度Tm,评估热稳定性。
3.6 生物信息前期工作
● 燕麦全基因组BLAST挖掘8个全新AsPgb,完成组织表达谱(根、叶、种子、颖壳等);
● ColabFold预测AsPgb三维结构,UCSF ChimeraX与已解析BvPgb1.2(PDB:7ZOS)骨架比对,RMSD<1 Å,整体折叠高度同源,但表面电荷、局部疏水存在差异。
四、实验结果
4.1 燕麦AsPgb基因与组织表达特征
1. 8个AsPgb分为3进化同源组,组内序列相似度>90%;
● OG0017468(AsPgb1.1–1.3):富集叶片、颖壳,种子表达低;
● OG0007273(AsPgb1.4–1.6):种子、根高表达,颖壳无转录;
● OG0012302(AsPgb3.1–3.2):全组织广谱高表达;
2. 所有AsPgb在根系均可检测到转录,与植物球蛋白参与根系氧化应激功能吻合。
| Orthogroup | Species ID | Seed n = 24 |
Glume n = 7 |
Spikelet n = 8 |
Leaf n = 11 |
Crown n = 4 |
Roots n = 8 |
| OG0017468 | AsPgb1.1 | 0 | 4 | 0 | 4 | 1 | 3 |
| AsPgb1.2 | 5 | 6 | 2 | 7 | 3 | 8 | |
| AsPgb1.3 | 0 | 7 | 1 | 10 | 3 | 8 | |
| Total: | 5/72 | 17/21 | 3/24 | 21/33 | 7/12 | 19/24 | |
| OG0007273 | AsPgb1.4 | 5 | 0 | 1 | 0 | 1 | 8 |
| AsPgb1.5 | 16 | 0 | 2 | 1 | 1 | 8 | |
| AsPgb1.6 | 16 | 0 | 3 | 0 | 1 | 8 | |
| Total: | 37/72 | 0/21 | 6/24 | 1/33 | 3/12 | 24/24 | |
| OG0012302 | AsPgb3.1 | 24 | 7 | 8 | 11 | 4 | 8 |
| AsPgb3.2 | 24 | 7 | 8 | 11 | 3 | 8 | |
| Total: | 48/48 | 14/14 | 16/16 | 22/22 | 7/8 | 16/16 |
表1. 不同同源组燕麦植物球蛋白(AsPgbs)在各组织中的检出情况
注:数值为检出阳性样本数量,n代表该组织总样本数;同源组合计为组内所有蛋白检出总数,分母为该组织最大可检出样本数。
4.2 结构比对结论
AsPgb1.1、1.5与BvPgb1.2主链RMSD仅0.859、0.945 Å,血红素结合口袋、保守半胱氨酸空间位置高度重合;但细微序列差异直接造成体内/体外表达行为完全不同。
| Protein | Alignment Score (A.U) |
Pruned Atom Pairs (#) |
RMSD (Å) |
| AsPgb 1.1 | 534.5 | 101 | 0.859 |
| AsPgb 1.5 | 603.7 | 130 | 0.945 |
表2 燕麦植物球蛋白预测结构与甜菜1型球蛋白标准晶体结构(BvPgb 1.2,PDB编号7ZOS)的叠合比对结果
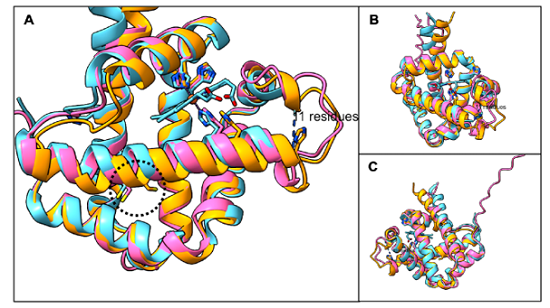
图3 AsPgb 1.1(橙色)、AsPgb 1.5(粉色)预测结构与BvPgb 1.2(蓝色)主链叠合图
4.3 微流控高通量筛选关键规律
1. 半胱氨酸C→A突变明显改变表达/纯化产率
● BvPgb C86A突变体:荧光表达总量高于野生型,纯化总收率与WT持平;SUMO标签强烈抑制野生型表达,但对C86A几乎无负面影响;
● AsPgb1.1 C70A:纯化收率高于野生;AsPgb1.5 C84A标签依赖性大幅改变;AsPgb3.1 C161A纯化效率大幅提升;
总体:消除保守巯基可减轻氧化聚集,多数Pgbs突变后可溶性、总产率提升,证明保守半胱氨酸是表达稳定性负向决定因子。
2. 氧化还原添加剂是Pgbs表达的关键调控因素
● GSSG(22%)、PDI/GSSG组合(26%)合计占优选反应体系48%,远高于分子伴侣DnaK(2%)、TRXB1(4%);
● 氧化微环境促进二硫键正确折叠、抑制巯基自发氧化聚集,是Pgbs稳定合成的核心条件;单纯还原型伴侣提升效果微弱;
● 金属辅因子混合液、Zn²⁺次之,可辅助血红素结合口袋折叠。
3. 可溶性标签具有蛋白/突变体特异性,小标签更优
● 表现较优标签集中在10 kDa以内:P17、CUSF、FH8;大标签SUMO/TRX/ZZ/SNUT普遍抑制Pgbs表达;
● 无标签体系占优选条件40%,说明标签并非必需,部分Pgbs去除融合标签后折叠更佳;3C蛋白酶添加组19%被选中,证实标签会阻碍部分Pgbs折叠;
● 物种差异:CUSF较适配BvPgb WT,P17较适配AsPgb1.5。
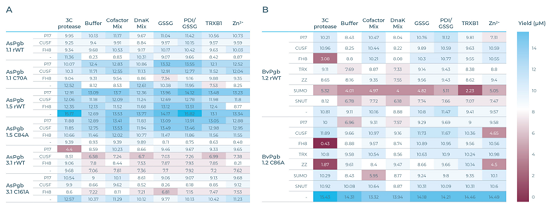
图4 各蛋白表达产量数据(μM),蓝色块产量>8 μM,红色块产量<8 μM
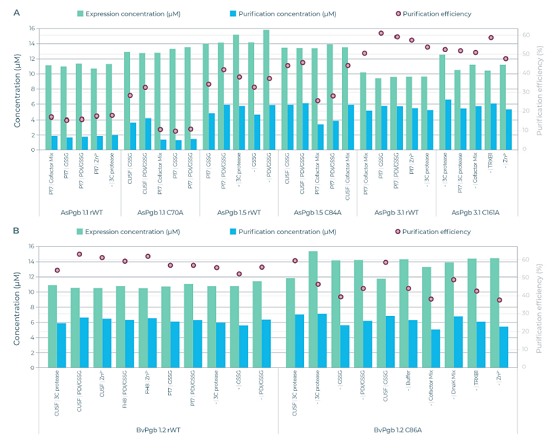
图5 各优选组蛋白表达产量(绿色)、纯化产量(蓝色)和纯化回收率(红色圆点)
4.4 放大表达与蛋白理化表征
1. 条件放大:AsPgb 1.5与BvPgb 1.2序列同源性高,AsPgb 1.5选用P17标签,BvPgb 1.2选用CUSF标签;均采用筛选效果优的PDI/GSSG反应液。放大纯化后获得P17-AsPgb1.5(0.69 mg/mL,22 μM)和CUSF-BvPgb1.2(0.77 mg/mL,20 μM),纯化后条带单一;
2. 质谱光度MP:两种蛋白均以单体为主(BvPgb单体占93%,AsPgb1.5单体72%),获得高纯度可溶性脱辅基apo-Pgb(无血红素结合);
3. 热稳定性nanoDSF(数据未展示):未开展完整热力学分析,BvPgb熔解温度Tm=55 ℃,高温易聚集;AsPgb1.5热转变信号不清晰,稳定性弱于甜菜同源蛋白,解释传统方案无法跨物种复用。
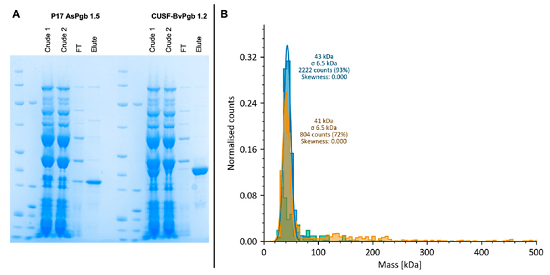
图6 P17-AsPgb 1.5(31 kDa)与CUSF-BvPgb 1.2(38 kDa)放大产物及寡聚状态表征
五、总结与展望
本研究依托微流控无细胞蛋白合成(DMF-CFPS)体系,实现了血红素结合球蛋白Pgbs高通量条件筛选,明确保守半胱氨酸易引发蛋白氧化聚集,将其突变为丙氨酸可大幅提升蛋白可溶性与纯化回收率;由氧化型谷胱甘肽GSSG与二硫键异构酶PDI组成的氧化环境更利于Pgbs正确折叠,且小分子亲水融合标签适配性好,大分子量标签会抑制蛋白折叠。不同物种同源Pgbs骨架虽保守,但序列差异导致理化性质与表达条件存在物种特异性,难以共用一套表达方案。
该体系可快速获得稳定可溶的脱辅基apo-Pgb蛋白,弥补了天然蛋白易聚集的短板,但目前尚未完成血红素嵌入及蛋白生物活性验证,突变位点改造维度较为有限。后续可添加GAPDH分子伴侣实现成熟全功能holo-Pgb制备,完善NO清除、抗氧化等功能检测,扩充氨基酸突变类型阐明巯基调控机理,并将这套高通量筛选系统拓展应用于膜蛋白、血红素酶、毒性重组蛋白等各类难表达蛋白的研发与机制解析,为血红素蛋白定向改造与应用开发提供技术支撑。
参考文献
Groth, L.; Bülow, L. Digital Microffuidics-Driven Cell-Free Protein Synthesis Platform Reveals Expression and Stability Determinants for Phytoglobins and Cysteine-to-Alanine Substituted Variants. Antioxidants 2025, 14, 1317.
拓展阅读
Nuclera无细胞蛋白表达筛选系统技术专栏(点击访问曼博生物官网查看)
本文基于Nuclera公开资料由中国抗体蛋白方案提供商上海曼博生物整理,用于科研信息分享、实验参考等。上海曼博生物可提供Nuclera eProtein Discovery无细胞蛋白表达筛选系统、数字微流控无细胞蛋白合成系统、DMF-CFPS高通量蛋白筛选、eGene可溶性标签试剂盒、难表达蛋白筛选、膜蛋白表达、血红素蛋白表达、蛋白稳定性筛选及蛋白纯化相关产品与技术支持,助力国内科研机构快速开展靶点验证、膜蛋白表达、AI蛋白设计和酶的定向进化以及酶工程等相关研究,如您需要欢迎咨询!