RNA 编辑的主要技术——CRISPR/Cas13 系统概述
2026-04-13 来源:本站 点击次数:118RNA 作为一种重要的生物大分子,参与细胞生命代谢的整个过程,其代谢异常会导致多种疾病的发生。 与相对稳定遗传的 DNA 不同,RNA 引起的改变是暂时的、非永久性的,但是 RNA 生物学的极度复杂性导致很难精确地将其靶向或编辑。目前靶向 RNA 的技术主要包括 RNAi 及 CRISPR-Cas13 技术等。
CRISPR/Cas13 系统是目前发现的能够靶向 RNA 的 CRISPR 系统,该系统的发现扩展了 CRISPR 系统在病毒干扰方面的应用。新发现的 Cas13 蛋白第四种亚型(Cas13d)的家族成员 CasRx 在体内外哺乳动物细胞的 RNA 裂解过程中显示出更强的特异性和更高的敲除效率,而且 CasRx 比之前发现的所有亚型都要小很多,更容易利用 AAV 载体实现体内递送,在 RNA 编辑方面具有很大的应用前景。
2016年,科学家发现了可以靶向RNA进行切割的CRISPR/Cas13系统,该系统由单一的效应蛋白Cas13和CRISPR RNA(crRNA)组装形成一个由crRNA引导的RNA靶向效应复合物。目前,Cas13家族共鉴定出四种亚型,包括Cas13a(又名C2c2)、Cas13b、Cas13c和Cas13d,四种Cas13亚型蛋白分子量均小于Cas9蛋白。
所有Cas13蛋白均具有两种不同的催化活性:
①由两个较高等的真核和原核核苷酸结合(Higher eukaryotes and prokaryotes nuceotide-binding,HEPN)结构域提供的RNase活性,这是靶RNA降解所必需的。Cas13 四个亚型同源性较低,且同源序列仅限于HEPN结构域位点,此外两个HEPN结构域中的单点突变会使CRISPR/Cas13系统对RNA的切割能力完全丧失;
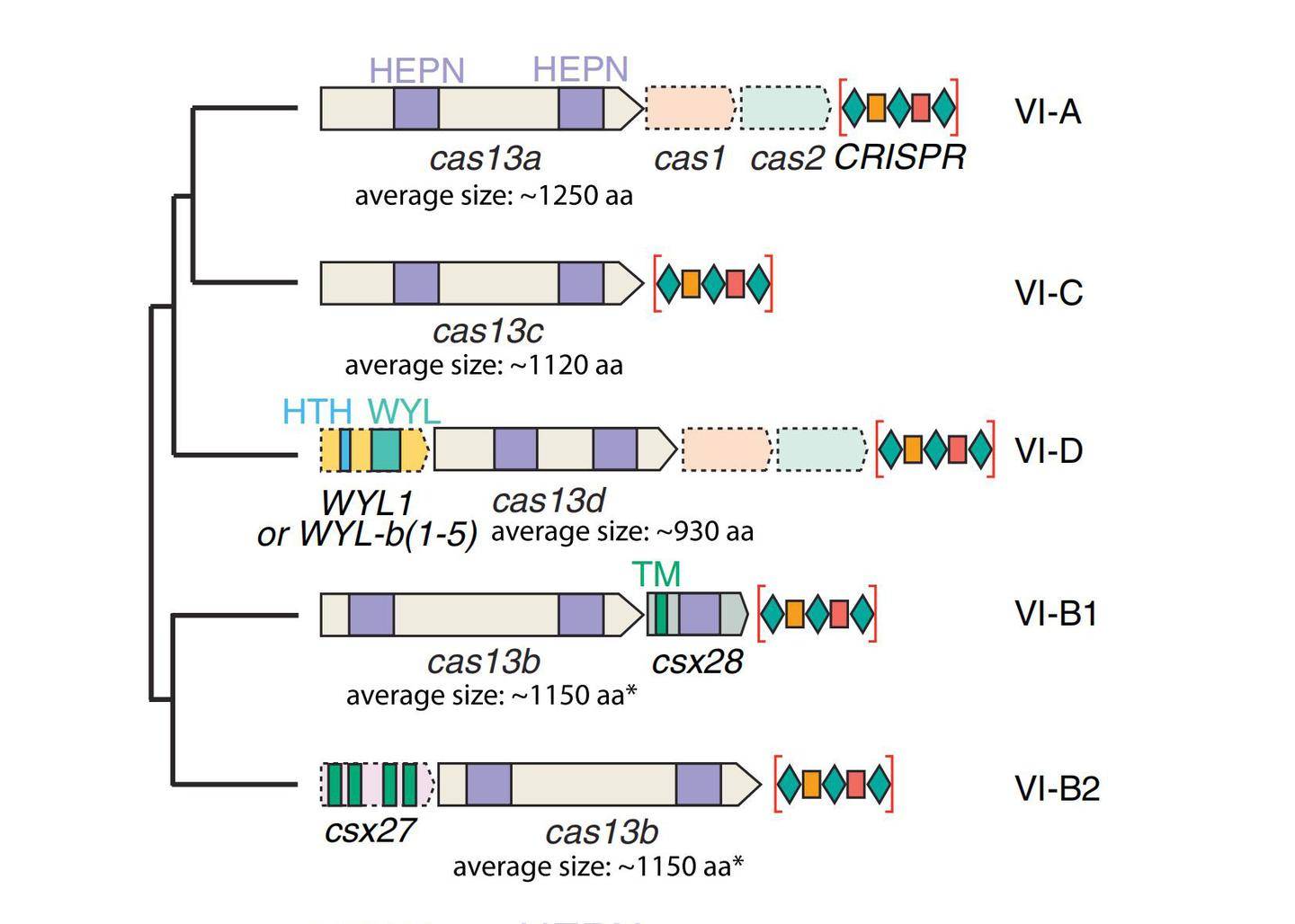 图1.CRISPR/Cas13系统介绍
图1.CRISPR/Cas13系统介绍
②催化pre-crRNA加工和形成成熟crRNA的RNase活性。
与CRISPR II类系统的其它核酸酶结构一致,Cas13a的整体结构是双叶的,N-末端结构域(N-terminal domain,NTD)和Helical-1结构域构成crRNA识别叶(REC lobe),HEPN1、Helical-2、HEPN2结构域形成核酸酶叶(NUC lobe)(图2)。CRISPR/Cas13b系统有两种酶,分别是VI-B1和VI-B2,它们之间的区别在于Cas13b转座子上携带的附属蛋白的基因型不同,VI-B1的附属蛋白是Csx28,而VI-B2的附属蛋白是Csx27。绝大多数Cas13d型系统还含有包含 WYL结构域的相关辅助蛋白,WYL结构域通常与原核防御系统有关(图1)。
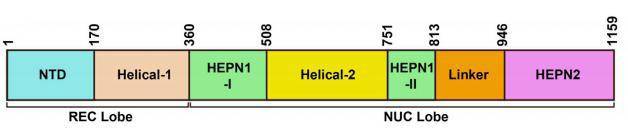 图2.Cas13a蛋白结构域示意图
图2.Cas13a蛋白结构域示意图
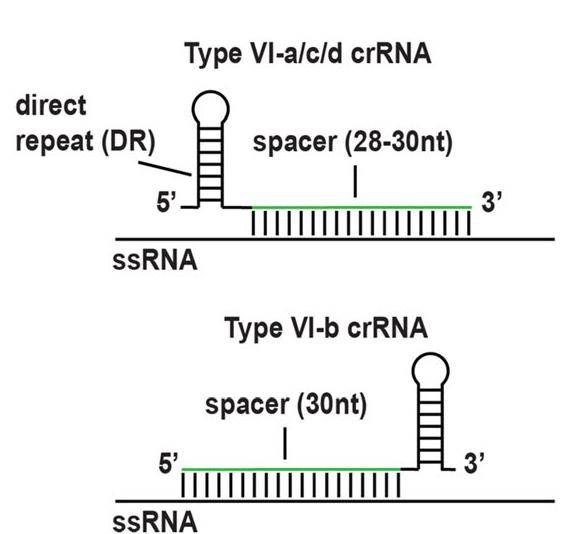
所有的Cas13蛋白酶都需要60- 66 nt的crRNA来确保靶向特异性。四种亚型crRNA都有一个促进与相应Cas13蛋白酶结合的直接重复(Direct repeat,DR)以及特异性靶向转录本的间隔序列。Cas13b的crRNA在3’端携带直接重复,而Cas13a、c和d的crRNA在5’端携带直接重复(图3)。