

紊乱的血流经Frizzled-4-β-catenin依赖性通路增加内皮炎症和通透性
2025-11-07 来源:本站 点击次数:690Disturbed flow increases endothelial inflammation and permeability via a Frizzled-4-β-catenin-dependent pathway
Keywords: Mechanotransduction, Inflammation, Orbital shaker, Atherosclerosis, Adherens junctions, Cytoskeleton
点击了解:流体剪切应力系统
内皮细胞持续受血流产生的壁剪切应力影响,其表面机械传感器通过机械信号通路响应这种应力。血管局部结构(如直形无分支区域与弯曲、分叉区域)决定了剪切应力的特性,进而导致内皮细胞功能差异。无分支区域与弯曲的高幅度、搏动性单轴剪切应力可维持细胞稳态,激活保护通路并促进相关蛋白表达。分叉区域则易引发内皮功能障碍,表现为通透性增加、eNOS 表达减少及促炎通路激活,与动脉粥样硬化易发性相关。
传统观点认为低时间平均壁剪切应力和高振荡剪切指数是引发内皮细胞功能障碍的关键,而近期研究更强调紊乱流或多向流的作用。虽然已知部分信号通路与流依赖性内皮细胞功能障碍相关,但紊乱流影响内皮细胞行为的具体机制仍不清楚。Wnt 信号通路不仅调控非血管机械响应细胞对机械力的反应,其在血管细胞中的作用也逐步明确。非经典 Wnt 配体参与血流依赖的内皮细胞极化,而在致动脉粥样硬化的血流环境中,Wnt 通路基因呈现富集状态,这提示该通路或许与血流介导的内皮细胞功能异常存在关联,且振荡剪切应力可通过增强 β- 连环蛋白信号激活 NF-κB。经典 Wnt 信号通路依赖 β- 连环蛋白的稳定化及核转运调控靶基因转录,其机制因多种配体、受体的组合作用而复杂。
点击了解:细胞力学刺激培养系统
基于此,伦敦帝国理工学院生物工程系的研究团队借助可模拟紊乱多向流的体外模型,探究了经典 Wnt 信号通路在内皮细胞对紊乱流的响应及功能障碍中的作用。研究成果发表在Journal of Cell Science期刊,题为“Disturbed flow increases endothelial inflammation and permeability via a Frizzled-4-β-catenin-dependent pathway”。

首先,由于之前有研究表明,Frizzled-4(FZD4)的 mRNA 水平对血流条件敏感,研究人员在不同血流条件下评估了FZD4 的表达水平。结果表明,与正常流和静态条件相比,紊乱流处理 24 小时可显著上调 FZD4 蛋白表达,且该效应能持续至 72 小时(图 1A),而正常流与静态条件下的 FZD4 表达无明显差异(图 1B)。此外,在生理状态下,猪主动脉弓内侧弯曲这一长期受紊乱流影响的区域,其 FZD4 表达也高于外侧弯曲的正常流区域(图 1C)。进一步支持了紊乱流对 FZD4 表达的调控作用,且提示该调控可能与内皮炎症信号传导相关。
为了探究 FZD4 是否介导长期紊乱流下的内皮功能障碍,研究人员通过 siRNA 敲低 FZD4进行了研究。发现紊乱流条件下,FZD4 敲低可显著降低促炎基因(SELE、MCP-1、VCAM1)的表达(图 1D),VCAM1 蛋白的表达也有所减少(图 1E)。而该效应仅存在于紊乱流环境中,在正常流下促炎基因本底表达较低,且不受 FZD4 敲低影响。这表明 FZD4 在紊乱流诱导的内皮促炎反应中起特异性介导作用。
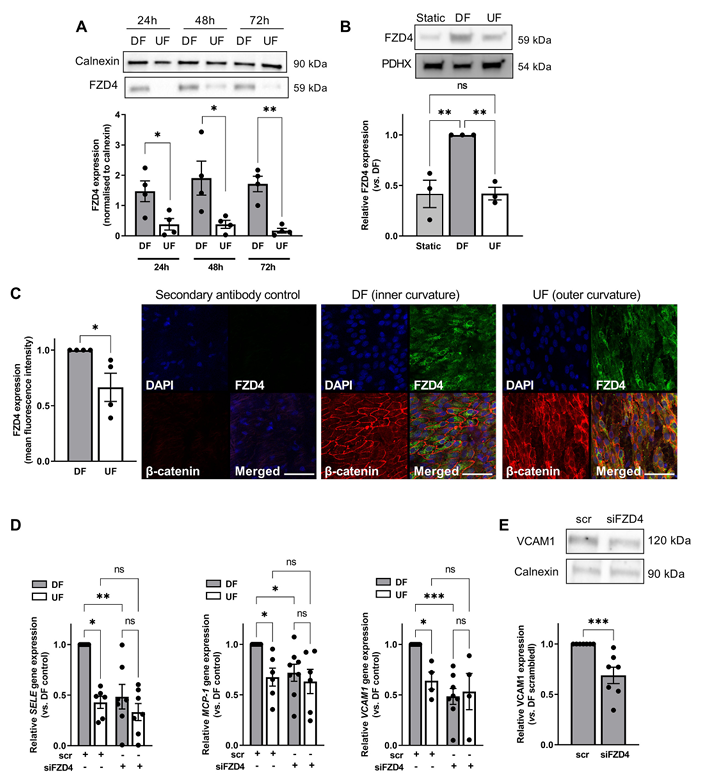
图1 在受到紊乱血流刺激的内皮细胞中,FZD4 的表达水平升高,并且其可调控促炎信号传导。
接着,研究人员进一步探究了紊乱流上调 FZD4 蛋白的机制。结果发现,紊乱流下 FZD4 基因表达低于正常流(图 2A),这提示其调控可能发生在转录后或翻译后水平。在紊乱流和正常流条件下 ,ZNRF3 的表达水平无明显差异(图 2B),但紊乱流可显著上调 RSPO-3 (一种可抑制 ZNRF3 的蛋白)的基因(图 2C)和蛋白表达(图 2D)(其他 RSPO 家族成员未检测到),而 RSPO-3 可抑制 ZNRF3 介导的 FZD4 溶酶体降解。实验证实,敲低 RSPO-3 会使紊乱流下的 FZD4 蛋白表达显著降低(图 2E、F),且会减少促炎基因 SELE 和 VCAM1 的表达(图 2G),与敲低 FZD4 的效应一致。这些数据表明,紊乱流下 FZD4 的表达受 RSPO-3 调控。

图2 在受到紊乱血流刺激的内皮细胞中,RSPO-3 的表达水平升高,并且其可调控 FZD4 的表达。
已知 FZD4 可激活依赖 β- 连环蛋白的经典 Wnt 通路,低剪切应力、振荡剪切应力或致动脉粥样硬化剪切应力可激活 β- 连环蛋白,但紊乱流是否具有相同效应尚不明确。为了验证 β- 连环蛋白在紊乱流下的作用,研究人员使用 iCRT5 抑制 β- 连环蛋白转录活性(图 3A),发现其可特异性降低紊乱流下促炎基因(SELE、MCP-1、VCAM1)的表达(图 3B),减少单核细胞与内皮细胞的黏附(图 3D),且对正常流条件无影响。此外,iCRT5 还能减弱 TNFα 诱导的促炎基因上调(图 3C)。这些结果表明,β- 连环蛋白在易致动脉粥样硬化血流引发的炎症激活中起调控作用,其效应与敲低 FZD4 或 RSPO-3 一致。
接下来,实验进一步证实 FZD4 与 β- 连环蛋白的关联。敲低 FZD4 会使紊乱流下 β- 连环蛋白的转录活性显著降低(图 3E)、蛋白水平下降(图 3F),而转录本水平不受影响(图 3G),且该效应仅存在于紊乱流环境。同时,敲低 RSPO-3 也会减少紊乱流下 β- 连环蛋白的表达(图 3F)。这些结果表明,FFZD4 依赖的 β- 连环蛋白激活至少在部分程度上介导了紊乱流的促炎效应。

图3 在受到紊乱血流刺激的内皮细胞中,β- 连环蛋白的活性增强,且这一过程依赖于 FZD4。
鉴于 Frizzled 受体常被 Wnt 配体激活,且 WNT5A 已知可促进内皮细胞炎症信号,研究人员探究了其在紊乱流响应中的作用。紊乱流会使内皮细胞中 WNT5A 的转录水平(图 4A)和蛋白水平(图 4B)表达显著增加。用 WNT5A siRNA 转染后,WNT5A 的表达降低了 35–40%(图 4C),且 SELE、MCP-1 和 VCAM1 的表达也相应减少(图 4D)。这些数据表明,WNT5A 可在紊乱流条件下促进内皮功能障碍。用 SFRP-1 阻断 WNT5A 与 Frizzled 受体的相互作用,可降低 β- 连环蛋白的转录活性(图 4E)。当细胞暴露于 SFRP-1 达 72 小时时,可显著降低 SELE 、MCP-1和VCAM1 的表达(图 4F)。这些结果表明,抑制 WNT5A 信号可减少紊乱流依赖的 β- 连环蛋白激活并减弱炎症信号。
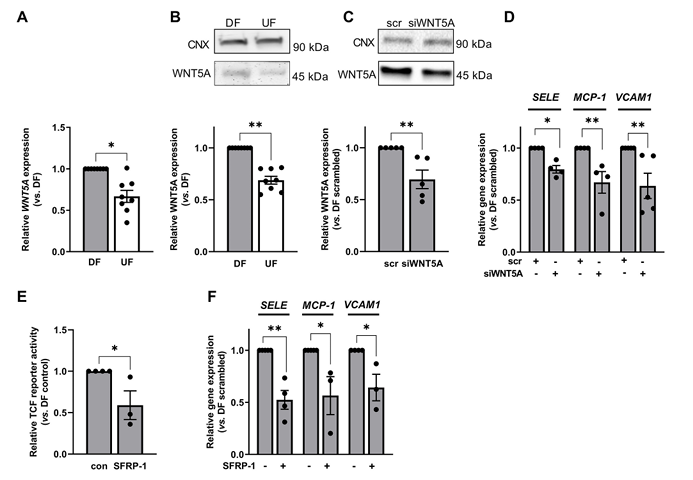
图4 在受到紊乱血流刺激的内皮细胞中,WNT5A 的表达水平升高。
然后,研究进一步探究了 FZD4 信号的下游机制。紊乱流会增加内皮细胞中 GSK3β 的 Ser9 磷酸化(该修饰可抑制 GSK3β,进而激活 β- 连环蛋白)(图 5A),而敲低 FZD4 或 RSPO-3 会减少这种磷酸化(图 5B),表明其通过影响 β- 连环蛋白降解复合体发挥作用。此外,用 IWR-1 稳定降解复合体可抑制 β- 连环蛋白转录活性,并减少SELE 和 MCP-1 的表达,与敲低 WNT5A、FZD4及抑制 β- 连环蛋白信号的效应一致(图 5C、D)。这些结果表明,紊乱流环境中,FZD4 信号通过 GSK3β 调控经典 Wnt 通路(依赖 β- 连环蛋白),进而影响内皮细胞功能 。
为了进一步探究 FZD4 激活 β- 连环蛋白的通路依赖性,研究人员评估了 LRP6 的激活情况(LRP6 激活是经典 Wnt 通路的标志)。结果显示,紊乱流与正常流条件下 LRP6 的磷酸化水平及总蛋白量无差异(图 5E、F) ,且用 DKK-1 阻断 LRP 与 Frizzled 受体的相互作用,对促炎基因表达(图 5G)及单核细胞黏附无影响,表明 FZD4 激活 β- 连环蛋白不依赖 LRP6。同时发现,紊乱流会上调内皮细胞中 Ryk 的表达(图 5H),敲低 Ryk 可减少紊乱流下 β- 连环蛋白的水平及 VCAM1 的表达(图 5I-K),表明 Ryk 可能作为替代组分参与 FZD4 介导的 β- 连环蛋白调控及炎症反应。

图5 紊乱血流可抑制 β- 连环蛋白降解复合体的功能,且这一过程依赖于 FZD4、RSPO-3 和 Ryk。
最后,由于抑制 β- 连环蛋白信号可改变紊乱流诱导的炎症激活,研究人员进而探究这种抑制作用是否同样会影响暴露于紊乱流的内皮细胞的屏障功能。使用 iCRT5 抑制 β- 连环蛋白转录活性后,可以显著降低紊乱流下 FITC - 亲和素的通透性(图 6A、B),对正常流无影响(图 6B)。同时,iCRT5 可改善紊乱流导致的连接结构紊乱,ZO-1 的表达增加,连接结构更有序(图 6C),并使细胞形态更规则(排列一致、更细长)(图 6C-E)。这些结果表明,紊乱流会增加内皮细胞的细胞旁通透性,而抑制 β- 连环蛋白活性可改善这一现象。

图6 在受到紊乱血流刺激的内皮细胞中,β- 连环蛋白的活性增强会增加细胞通透性,并改变细胞骨架及连接结构的组织方式。
研究进一步探究了 β- 连环蛋白抑制对细胞结构的影响。iCRT5 处理可减少紊乱流下内皮细胞的应力纤维数量,增加连接部皮质肌动蛋白染色(与连接稳定、通透性降低相关)(图 6D),并使纽蛋白从粘着斑更多地定位到连接部周围(图 6E)。而敲低 FZD4 也会引发类似的细胞形态、连接结构、细胞骨架及纽蛋白定位变化(图 7A-C)。敲低 WNT5A,会引发细胞形态(伸长)、细胞骨架(应力纤维减少、皮质肌动蛋白增加)及连接结构变化(图 7D)。同时,抑制 β- 连环蛋白信号通过上调 PECAM-1、JAM-A、JAM-C(JAM3)、ZO-1(TJP1)和 CLDN5 的表达(图 7E) ,稳定细胞间连接,从而增强内皮屏障功能。这些数据表明,抑制 β- 连环蛋白信号可增加紊乱流暴露的内皮细胞中连接蛋白的表达并稳定细胞间连接。
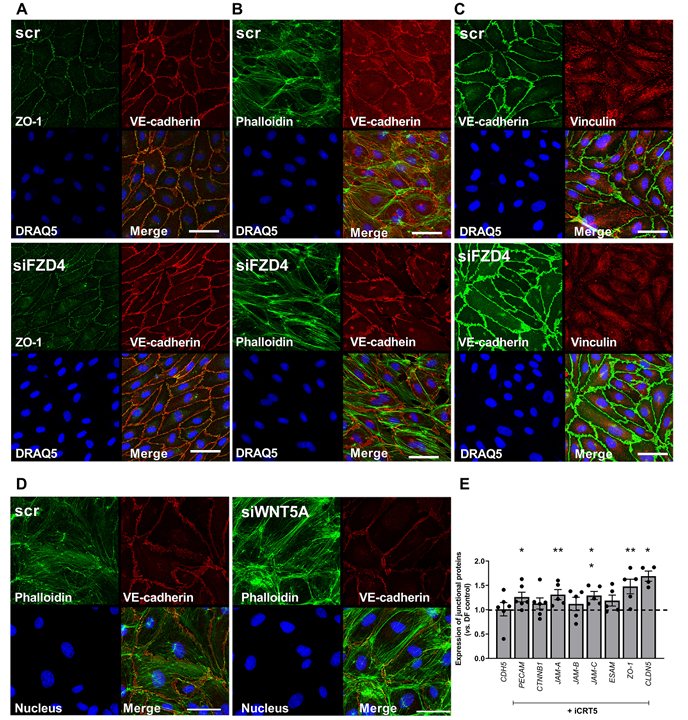
图7 在受到紊乱血流刺激的内皮细胞中,敲低 FZD4 或 WNT5A 会改变细胞骨架和纽蛋白的组织方式。

图8 紊乱血流依赖性的 FZD4-β- 连环蛋白信号通路的推测机制。
总之,该研究表明,紊乱流会通过上调 RSPO-3 来增加 FZD4 的表达,FZD4 进而不依赖 LRP 信号激活 β- 连环蛋白的转录活性,最终导致内皮细胞的炎症激活和屏障功能破坏(图 8)。这一通路的发现揭示了紊乱流诱导内皮功能障碍的新机制。
参考文献:Rickman M, Ghim M, Pang K, von Huelsen Rocha AC, Drudi EM, Sureda-Vives M, Ayoub N, Tajadura-Ortega V, George SJ, Weinberg PD, Warboys CM. Disturbed flow increases endothelial inflammation and permeability via a Frizzled-4-β-catenin-dependent pathway. J Cell Sci. 2023 Mar 15;136(6):jcs260449. doi: 10.1242/jcs.260449. Epub 2023 Mar 24. PMID: 36846872; PMCID: PMC10112981.
原文链接:https://pubmed.ncbi.nlm.nih.gov/36846872/
图片来源:所有图片均来源于参考文献
小编旨在分享、学习、交流生物科学等领域的研究进展。如有侵权或引文不当请联系小编修正。如有任何的想法以及建议,欢迎联系小编。感谢各位的浏览以及关注!进入官网www.naturethink.com或关注“Naturethink”公众号,了解更多相关内容。
点击了解:流体剪切应力系统
点击了解:细胞力学刺激培养系统