

依据颜色和光线的物理测量肉类颜色指南
2025-11-14 来源:本站 点击次数:470
肉类颜色测量指南
颜色和光线的物理
A. 导言
感知物体并识别其颜色的过程涉及多重因素:物体本身、其周围环境,以及负责感知并转化刺激信号为颜色感知的检测器。要实现颜色感知,必须配备具备这种功能的检测器。这类检测器可以是人眼,也可以是色度计、分光光度计等专业仪器。
人类的感官反应与色彩识别,需要眼睛和大脑协同工作来感知并处理外界刺激以辨别颜色。眼睛由角膜、瞳孔、虹膜和晶状体构成,共同形成眼球的前房。晶状体将前房与后房(玻璃体)分隔开来,后房内含有视网膜和视神经。眼睛的工作原理与相机类似:光线穿过瞳孔,经晶状体聚焦后投射到视网膜上。虹膜则像相机快门一样运作,在光线较暗时开启以增加进光量,在强光环境下收缩以限制光线进入。
视网膜是感知光线的细胞器,其感光细胞分为视杆细胞和视锥细胞。视杆细胞不敏感于颜色,但能感知从黑到灰再到白的视觉光感;而视锥细胞则对可见光的颜色敏感(图3.1)。根据对光谱的峰值响应范围,视锥细胞可分为蓝、绿、红三种类型。因此,当光线进入眼睛时,视杆细胞负责检测明暗刺激,视锥细胞则负责识别蓝、绿、红三种光谱。这种对蓝、绿、红光谱的感知能力被称为眼睛的三色视觉功能。这些视觉信息随后通过视神经传递至大脑,在大脑中经过处理后,最终形成对物体的视觉感知。因此,眼睛与大脑的复杂互动是颜色感知的形成基础。这一过程受多种因素影响,可能扭曲颜色感知,尤其是颜色的检测方式及其处理机制。要确定颜色,需要一种能够捕捉这些信息的检测器。
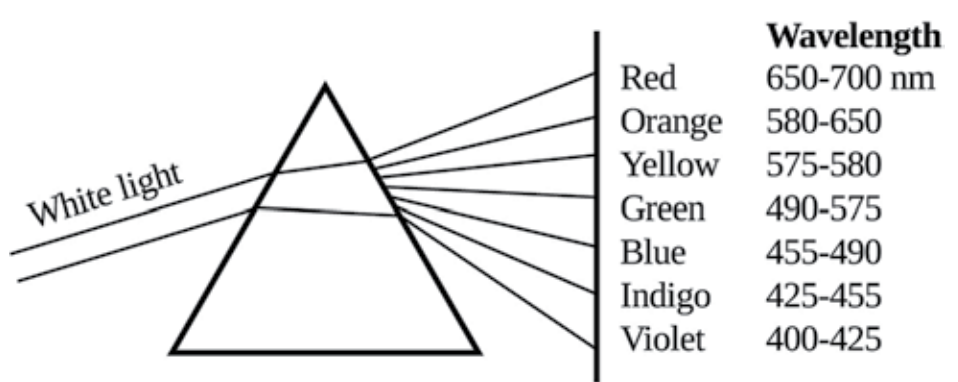 图3.1. 白色光通过棱镜分解成其组成成分。在光被分解的点上放置另一个棱镜,将再现白色光。由圭尔夫大学Shai Barbut博士提供。
图3.1. 白色光通过棱镜分解成其组成成分。在光被分解的点上放置另一个棱镜,将再现白色光。由圭尔夫大学Shai Barbut博士提供。
信息获取是必要的。然而,并非所有眼睛都具备同等的光感检测和色彩感知能力。以红绿色盲为例,部分人群存在这种先天缺陷。虽然光谱本身能产生色彩感知,但眼睛无法有效识别,大脑也无法正确处理这些视觉信号。因此,任何色彩测量都必须确保检测器官正常运作——就像人类色彩感知系统那样——这样才能准确检测红绿色盲等色彩识别障碍。目前网络上已有专门用于检测色盲的标准化测试表。
需要说明的是,人眼或其他机械装置并不会“看见”颜色,它们只是捕捉 来自物体(如肉类)的反射光波长(图3 . 2), 而人眼则将这些感官信息 传递给大脑进行解析。
肉类或其他物体的颜色,实际上是光线、视觉系统(即感光器官)与被观察物体相互作用的结果。光线对色彩感知至关重要——没有光线,就既没有颜色也谈不上视觉。可见光是电磁波谱的一部分 ,该波谱由不同波长的能量定义 ,涵盖广播波、雷达波、红外线、紫外线、X射线、伽马射线及宇宙射线。但人类只能感知波长在390至750纳米之间的可见光。在这个狭窄的电磁波谱范围内,人眼具备将波长区分为不同颜色的能力,而大脑则拥有将这些波长归类为特定颜色的处理能力。例如,红色通常对应波长约650至700纳米的光线。
绿色对应波长范围约490-575纳米,蓝色对应455-490纳米(图3.1)。
有趣的是,其他动物,比如蜜蜂,可以在紫外范围内看到,蝙蝠可以感知电磁辐射在超声波范围内的光谱。
要检测颜色 , 光必须从被观察物体反射回来并返回眼睛 。要产生颜色,照射物体的光必须具有光谱范围
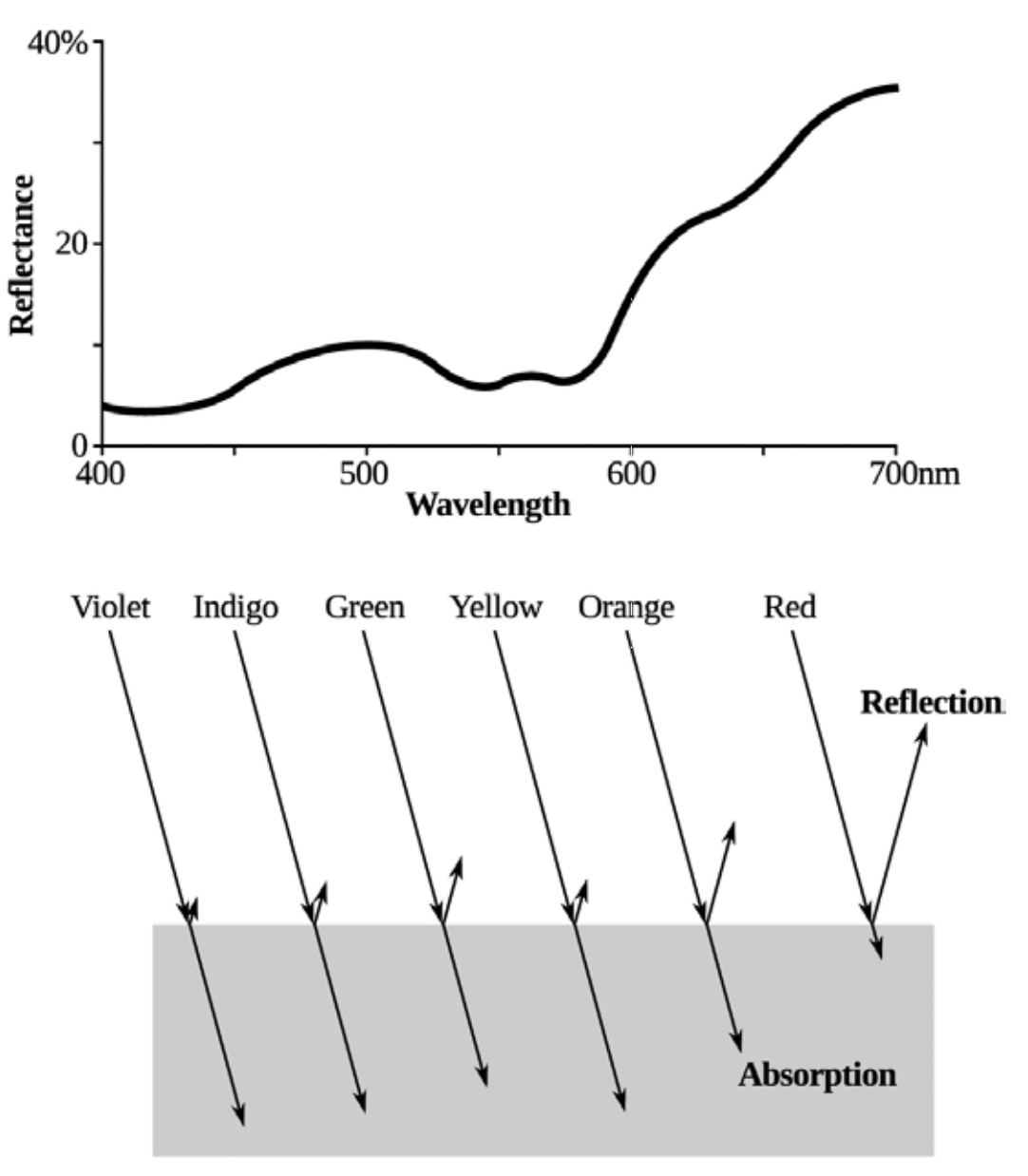 图3.2. 牛肉切片的光谱反射率(上图)和不同波长的相对反射率(下图)。请注意,观察者只能看到从表面反射的波长/颜色,而看不到被表面/肉吸收的波长。由圭尔夫大学Shai Barbut博士提供。
图3.2. 牛肉切片的光谱反射率(上图)和不同波长的相对反射率(下图)。请注意,观察者只能看到从表面反射的波长/颜色,而看不到被表面/肉吸收的波长。由圭尔夫大学Shai Barbut博士提供。
人眼能够感知并由大脑解析为颜色的对应波长,取决于光的反射特性。因此,当完整的可见光谱入射时,理论上可以产生无限种颜色。当光线照射到物体表面时,会经历吸收、反射或散射三种过程。被物体吸收的光波长无法被肉眼察觉,因为这些光线被物体(如肉类)吸收(见图3.2)。而反射回来的光线则会被眼球捕获,传递至大脑进行处理。由于人眼具有三色视觉 , 大脑会解析 蓝、绿、红三种颜色的光强度,最终将这些信息整合为我们所感知的颜色。
因此,要准确判断肉类颜色,光源必须包含能有效反射肉表面的波长,否则肉色将无法被肉眼或仪器检测器识别。在感官和仪器检测肉类时,光源必须标准化。总体而言,要让人类准确观察物体的真实颜色,应使用平衡光源。举例来说,使用白炽灯(图3 . 2和3 . 3)会使新鲜肉类呈现更鲜艳的红色,而 红外辐射输出较低的5000K荧
光灯(图3 . 3)则会呈现较暗的红色。
除了光检测、色彩生成与感知涉及的物理原理外,多种物理条件还会影响肉品色泽。本文重点探讨同一肉块在不同条件下如何产生差异化的色彩感知,而非深入分析肉品本身的色素构成或化学成分。 影响色泽的关键因素包括:光源特性 、观察者个体差异、肉块尺寸差异、表面光滑度(例如使用锋利刀具与钝刀切割)、背景环境差异以及观察角度差异。
肉类反射的光线波长决定了颜色的感知效果 , 因此光源 对颜色的呈现与感知起着关键作用。 光源 有多种类型 , 包括自然光 、 荧光灯 、 白炽灯 等,即便同一种光源,不同设备的发光特性也可能大相径庭。每种光源都具有独特的光谱组成。图3.3展示了两种光源的光输出对比。
所谓平衡光源,是指能均匀输出不同波长光线的光源(例如自然光)。正因如此,肉类在零售展示柜中可能呈现理想色泽,但在商店照明下(例如,许多商店在展示冷藏柜中使用荧光灯,因为这种灯泡发热量极低且比白炽灯更节能——后者会损失超过70%的能量为热量)反而会失去诱人外观。因此,在选择照明设备时
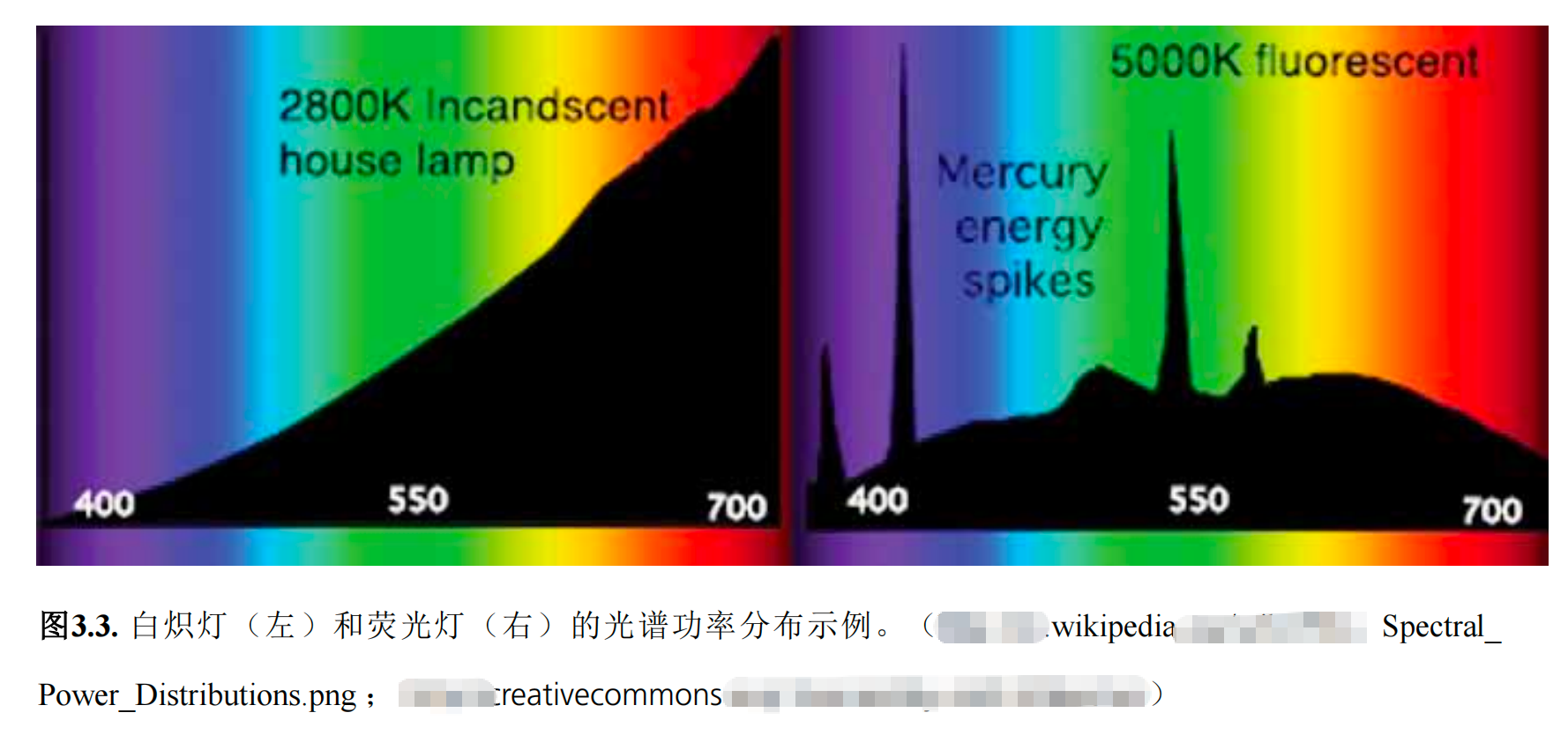
要准确比较肉品颜色,必须保持光源类型和照明条件恒定。肉品观察常用豪华暖白荧光灯作为光源。除了光源类型,光照强度对颜色感知同样关键——肉品观察时既不能过亮也不能过暗。比较肉品颜色时通常采用约1630勒克斯的光照强度。图3 . 4展示了三块新鲜肉品在冷白荧光灯照射下的实际反射波长。这些光谱正是消费者肉眼所感知和评估的。该灯泡在靛蓝、绿色和橙色区域有明显峰值(与图3.3中的5000K白炽灯相似),但红色区域亮度极低;因此消费者小组将牛肉、猪肉和鸡肉切块判定为棕色。相比之下,当使用白炽灯(反射光谱与图3 . 2所示及图3 . 4中2800K白炽灯 相似)照射时,肉品呈现的色泽则截然不同。3 ), 消费者小组的评分 更多是粉色/红色。
观察者差异是影响颜色感知的另一个关键因素。每个人的视觉敏感度在颜色感知上存在细微差异,这可能是所有影响颜色感知的因素中最难 控制的。通过颜色视觉筛查测试,可以筛选出能够准确辨别肉品颜色差异的评审专家(参见色盲测试表;Wiegand和Waloszek,2003)。需要特别注意的是,若未对屏幕进行完整校准,使用电脑呈现这些测试表时可能会出现偏差。
肉块的大小差异也会影响颜色的感知,因为反射到眼睛的光线的数量。对于较大的肉块,反射到眼睛的光线更多,颜色通常被感知为更明亮和更鲜艳。
背景差异同样会影响色彩感知。当在明亮背景前观察切片时,颜色往往显得暗淡无光;而在深色背景前观察时,切片则显得更加鲜亮。为确保色彩对比的准确性,需统一背景标准。
在肉类摄影中,背景色尤为重要——浅色背景容易造成颜色暗淡或苍白的视觉错觉,而深色背景则能更好地呈现肉质的鲜艳色泽。
此外,观察切面的角度和光源入射角都会影响颜色感知。当出现光泽时尤其重要,这可能妨碍观察样品。对于虹彩现象等条件,观察者的入射角会决定该现象的可见性。应避免背光照明,建议使用顶光。设置灯光时,需用光度计对亮度进行标准化测量。
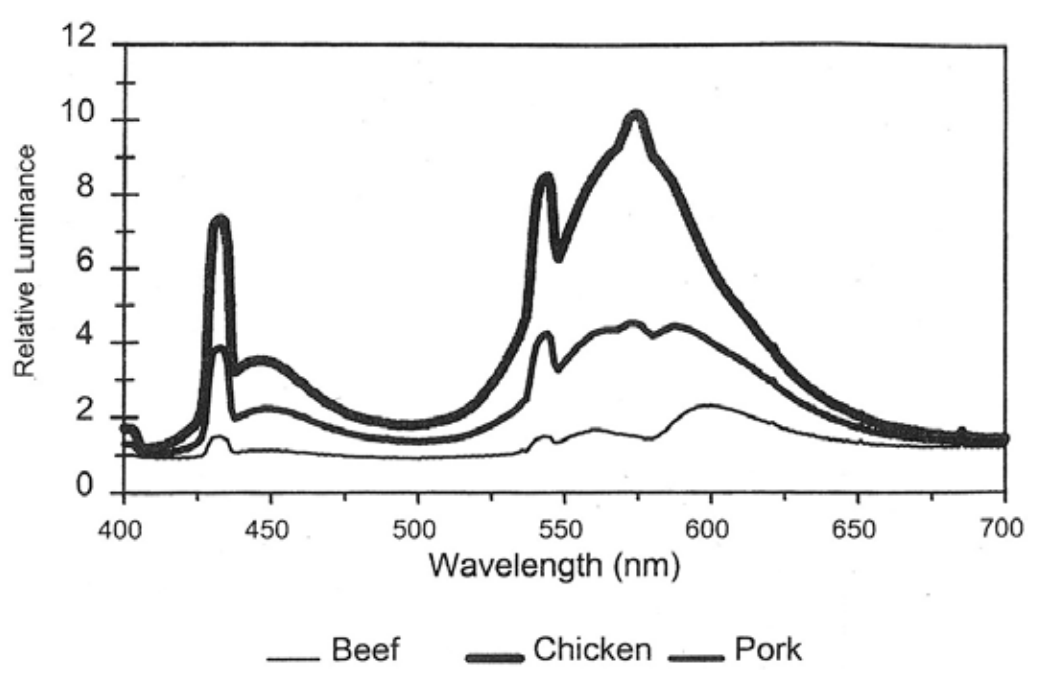 图3.4. 新鲜肉块在冷白荧光灯下呈现的相对亮度对比。需注意该灯具在靛蓝、绿色和橙色区域存在明显色温峰值。由于光源在可见光谱末端的亮度较弱,导致牛肉和猪肉切块的红色调难以清晰呈现。本文转载自《肉类科学》第59卷第2期S·巴布特(S. Barbut)发表的《照明光源对鲜肉切块外观的影响》(2001年,第187-191页),经爱思唯尔出版社授权使用。
图3.4. 新鲜肉块在冷白荧光灯下呈现的相对亮度对比。需注意该灯具在靛蓝、绿色和橙色区域存在明显色温峰值。由于光源在可见光谱末端的亮度较弱,导致牛肉和猪肉切块的红色调难以清晰呈现。本文转载自《肉类科学》第59卷第2期S·巴布特(S. Barbut)发表的《照明光源对鲜肉切块外观的影响》(2001年,第187-191页),经爱思唯尔出版社授权使用。
B. 肉类的色彩感知
当光线照射到肉品表面并反射回检测器(眼睛或仪器)时,处理器(大脑或微处理器)就会解析颜色信息。颜色传达其实颇具挑战性。为此,人们开发了专门工具来帮助解读色彩语言。美国艺术家A·H·芒塞尔发明的芒塞尔系统,通过色卡进行颜色混合与比对。
1931年,国际照明委员会(CIE)制定了XYZ三刺激值(图3.5),并于1976年推出CIE L*a*b*色空间(图3.6)。之所以开发CIE L*a*b*系统,是因为XYZ色度测量法中各颜色间的距离,并不能准确反映人眼感知到的颜色差异。例如,绿色与黄绿色的色差相对较大,而蓝色与红色的色差却微乎其微。为解决这个问题,CIE在1976年推出了三维Lab色空间(又称CIELAB 色空间)。在这个系统中,人眼感知到的颜色差异,与色度测量法得出的距离完整对应。a 轴从绿色(-a)到红色(+ a),b 轴从蓝色(-b)到黄色(+ b)。亮度(L)从三维模型的底部到顶部增加(图3.6)。
在报告研究用色度计值时,作者必须注明使用的是CIEL*a*b*值还是CIELab值。(星号的存在或缺失反映了 这些值的计算方式的细微数学差异。)可感知的颜色具有色相、明度和饱和度三个特性。色相是 我们用语言描述颜色时 所用的词汇(红色 、 黄色 、 绿色 、 蓝色 、等等)。色相由肉品表面反射回检测器的特定波长决定。明度描述颜色的明暗程度 ,而饱和度则反映颜色的鲜艳或暗淡程度。目前已有多种标准化方法用于颜色的测量与描述。
XYZ三刺激值及其对应的Yxy色空间为颜色描述提供了范式6(图3.5)。基于此,国际电工委员会(CIE)开发了x、y色度图。该图示将无色系(如浅色或暗淡色,饱和度较低)置于图中心,色度值则向图外围递增
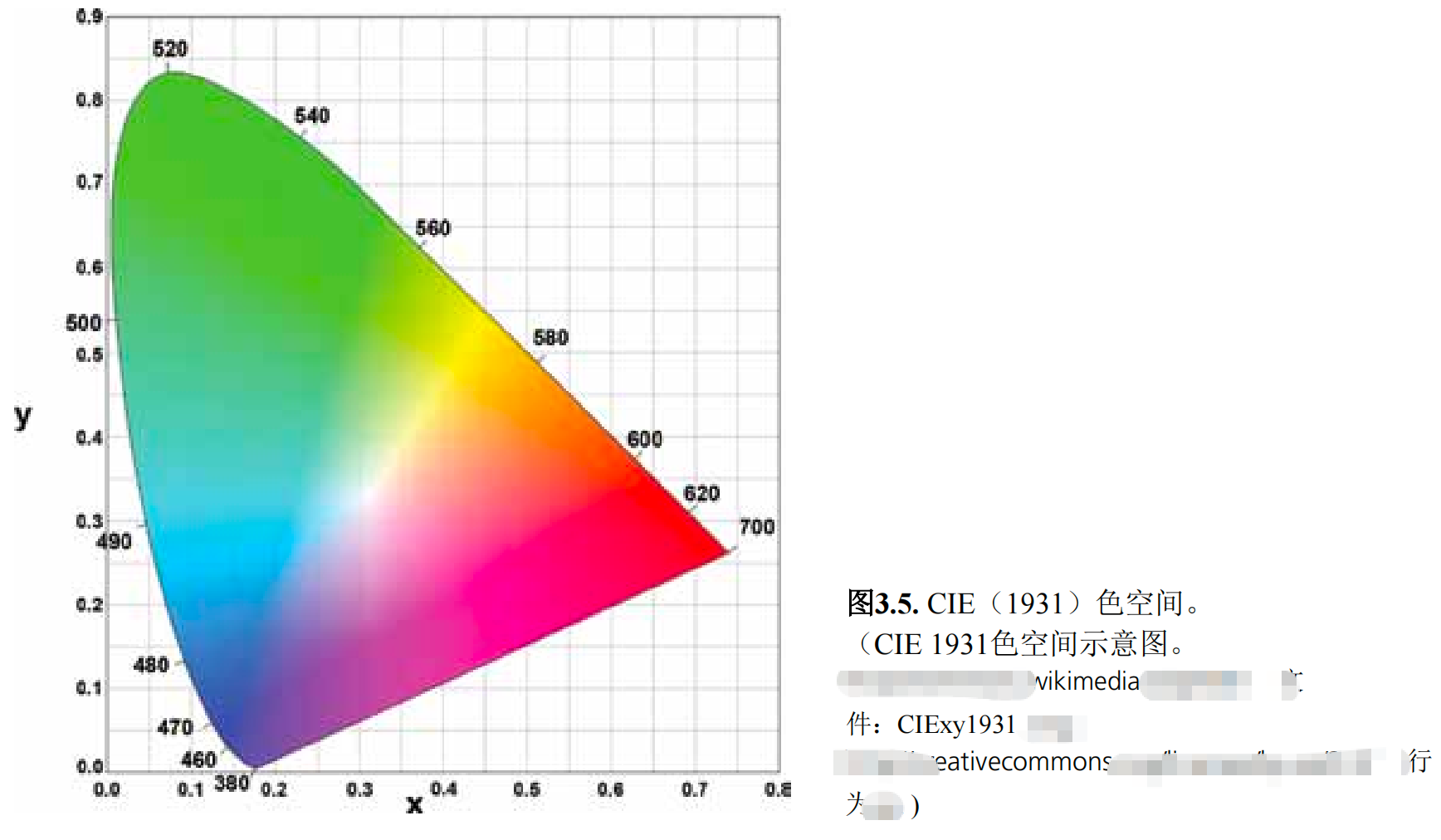
(色彩更鲜艳、饱和度更高)。在色环外围,红色、绿色和蓝色作为原色,对应着这些颜色的可见光波长。色度图通过坐标系绘制x轴和y轴的色值,从而确定颜色的色调和饱和度。
CIE L*a*b*色空间的后续发展使得颜色能够以三维空间形式呈现(图3.6)。由于人眼对蓝、绿、红三种颜色的视觉反应特性,相关计算将这些反应转化为L*、a*和b*三个数值。当这三个数值结合时,便构成了三维色空间。在该色空间中,a*值位于X轴,b*值位于Y轴,L*值位于Z轴(图3.6)。色空间中心为中性灰。沿a*轴方向,正值表示红色,负值表示 绿
色(红色范围 为+ 60至绿色范围 的-60)。沿Y轴 ,正b *表示黄色,负b*表示蓝色(红色范围为+ 60至蓝色范围的-60)。第三个维度L*的数值范围为100代表白色,0代表黑色(图3.6)。在此色空间中,通过绘制a*和b*的数值可确定肉样颜色或色调(图3.7)。通过L*值可测定样品的明度或暗度。因此,
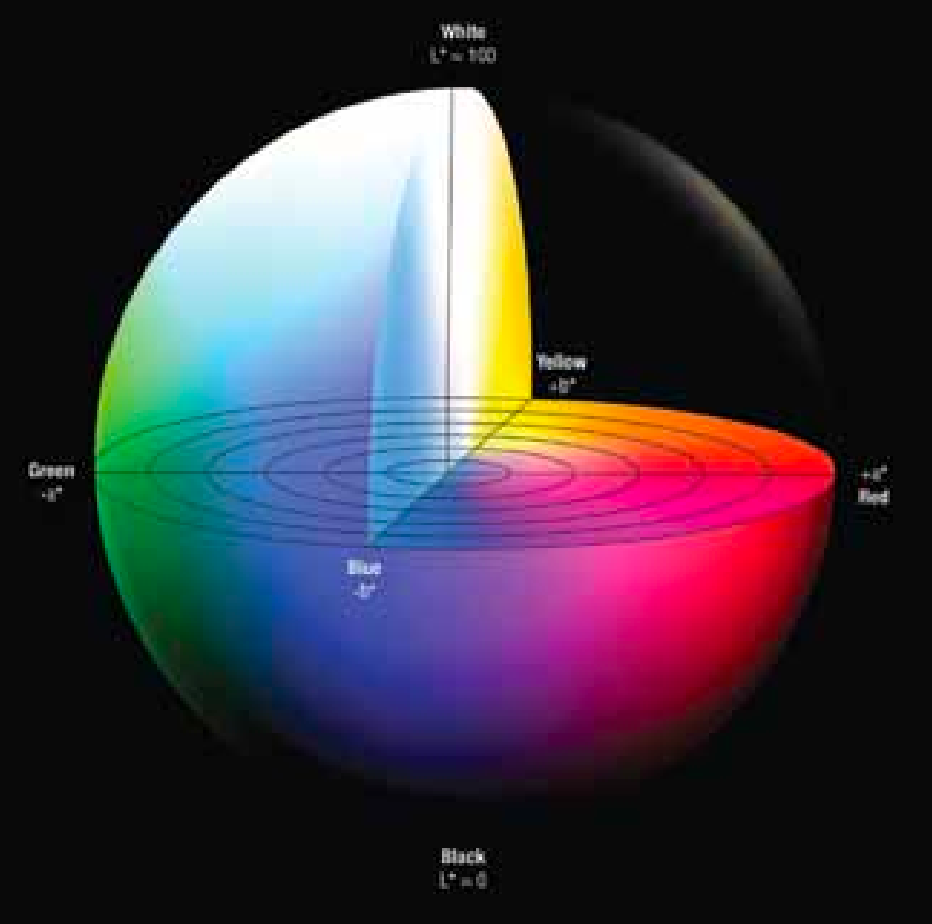 图3.6. CIE L*a*b*色空间的彩色实线表示。 图像由Konica Minolta Sensing Americas提供。
图3.6. CIE L*a*b*色空间的彩色实线表示。 图像由Konica Minolta Sensing Americas提供。
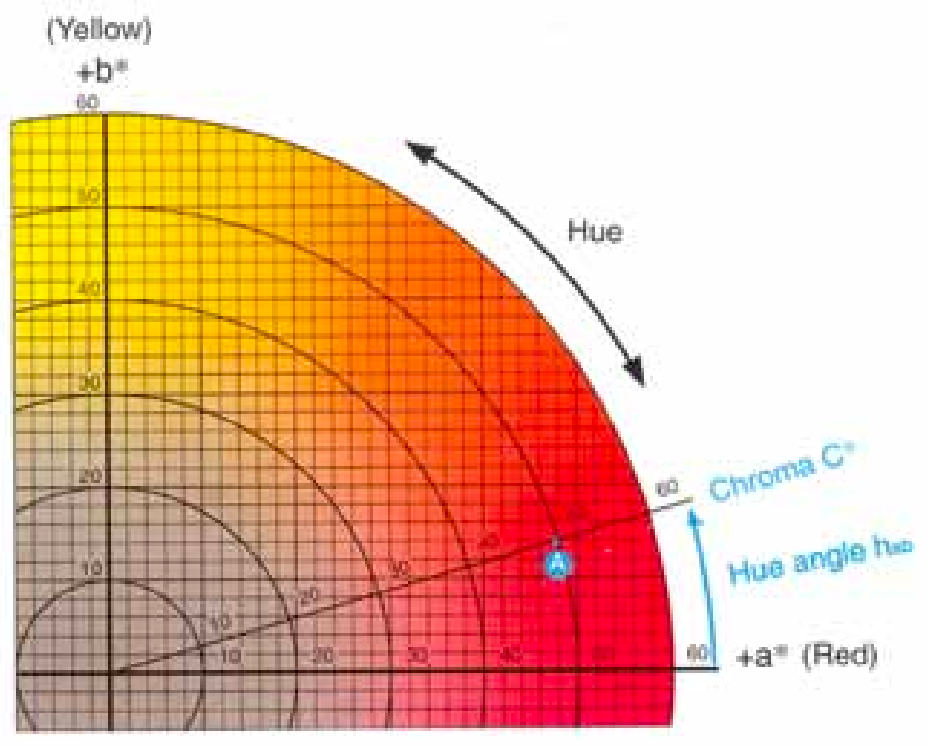 图3.7. 色度图中色相角与色度C*(饱和度指数)的示意图。点A是CIEa*(47.63)和CIEb(14.12)的图。
图3.7. 色度图中色相角与色度C*(饱和度指数)的示意图。点A是CIEa*(47.63)和CIEb(14.12)的图。
通过三角函数计算,可以确定样品偏离X轴的入射角,进而计算出样品的色相角(即颜色),同时通过计算样品与XYZ坐标系原点的距离来确定其饱和度或鲜艳度。色相角的计算。图片由Konica Minolta Sensing Americas提供。
公式为h=arctan(b*/a*)。例如,当样品的CIEb*值为14.12,CIEa*值为47.63 6时,其色相角为16.51度。因此,a*和b*值点与对应角度的坐标图,能够完整呈现样品的颜色特征。由于颜色在色域边缘的鲜艳度逐渐增强,a*和b*坐标点离原点越远,颜色的鲜艳度就越高。此外,色度(饱和度指数)也可通过公式(a*+b*)计算得出。例如,当a*值为47.63,b*值为14.12时,12 ,色度(饱和度指数)将为49. 68。有了这些数据 , 可以计算和比较色差 。这些计算将在以下章节中进一步讨论。
C. 光的物理性质与仪器色彩测量
仪器套装主要分为两大类用于颜色测量:色度计和分光光度计。这两类设备都配备专用光源和特定照明条件(例如A、C或D型光源 )。可选用不同类型的光源(如钨丝灯或氘灯).在选择光源时,光源的光谱特性、灯泡成本等因素都会成为重要考量。不同仪器在测量反射光的方式上存在差异。三刺激法使用光源照射样品,光线经红、绿、蓝滤光片后投射到光电探测器(图3.8)。微处理器可将反射值转换为XYZ或CIE L*a*b*值。分光光度计则通过扫描(通过单色仪)或光电二极管阵列同步读取(图3.9)的方式获取反射波长。这些数据传输至微处理器后,既可呈现为反射光谱,也可转换为如图3.9所示的XYZ值或CIE L*a*b*值 。
部分反射光谱分光光度计采用衍射光栅技术,通过扫描表面反射的波长(即颜色)进行检测,而另一些则通过检测反射光的特定波段来工作。
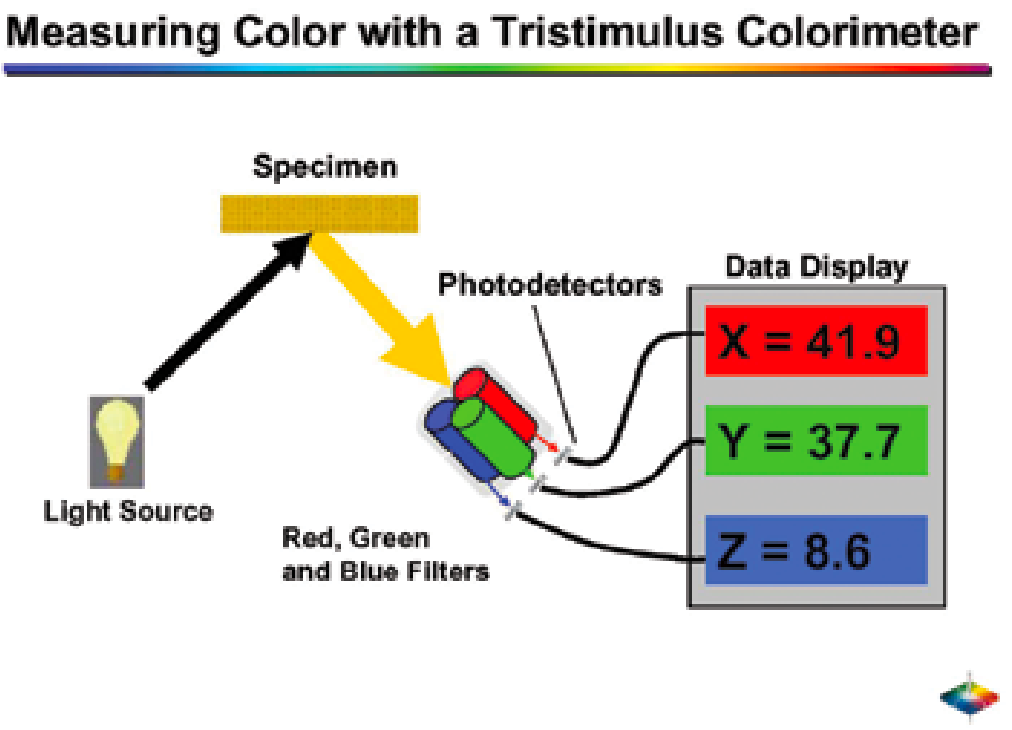 图3.8. 三刺激色度计示意图。 图片由HunterLab提供。
图3.8. 三刺激色度计示意图。 图片由HunterLab提供。
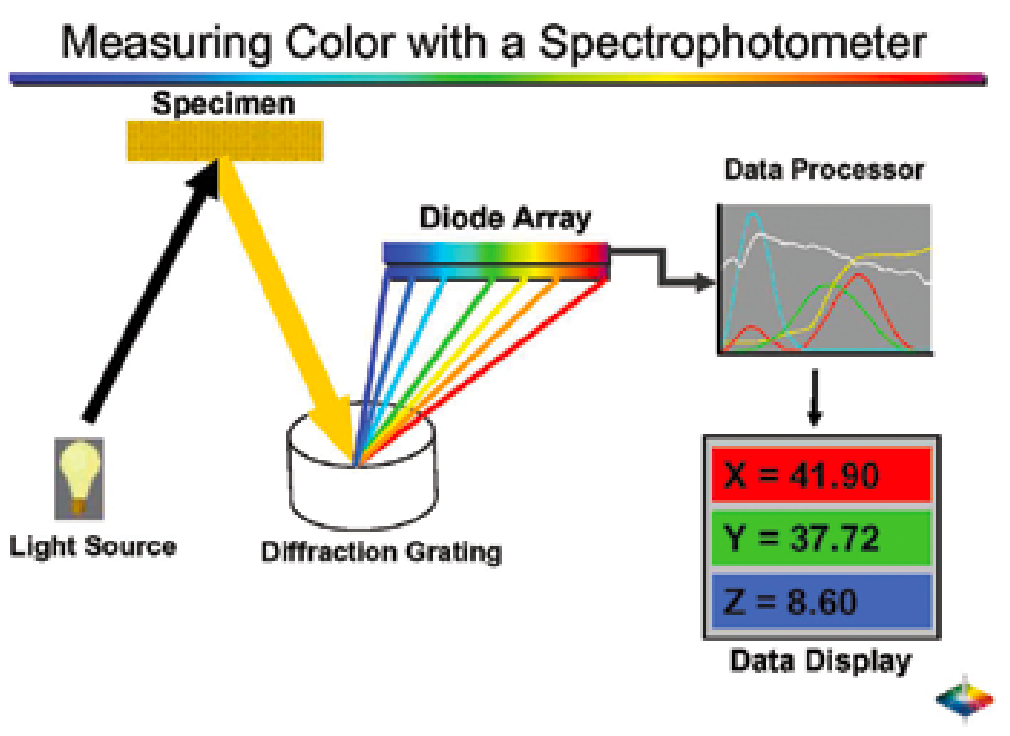 图3.9. 配有光电二极管阵列的分光光度计测量颜色的示意图。 图片由HunterLab 提供。
图3.9. 配有光电二极管阵列的分光光度计测量颜色的示意图。 图片由HunterLab 提供。
通过使用光电二极管阵列(例如一种能根据工作模式将光转换为电流或电压的光电探测器)
实现这一功能。衍射光栅本质上是一种带有大量平行且间距紧密的狭缝的固体板,或是带有众多平行反射凹槽的板。值得注意的是,肉表面本身也能充当衍射光栅。完好肉块表面呈现的虹彩效应,源于肌纤维内部肌原纤维的高度有序结构。当表面被切割时,会形成类似反射凹槽的结构。在这种情况下,入射光会被衍射(类似棱镜的原理),形成多种色调。
然而,分光光度计的衍射光栅能比棱镜更有效地分离不同颜色的光,其色散效应更为显著。即便是单一波长的光也能被进一步衍射。光电二极管阵列的设计初衷是同步测量不同波长的光信号。部分光电二极管阵列的分辨率可能仅有2至10纳米,因此当需要检测非常尖锐的反射峰或谷时,扫描反射光谱仪可能是更优选择。但随着光电二极管阵列技术的不断进步,这种优势可能会逐渐消失。相较于三刺激测量法,这种高分辨率特性在色素分析中更为关键。需要特别注意,扫描反射光谱仪在目标可见光波长范围内采集反射数据的速度,远低于二极管阵列的采集速率。
此外,需要特别注意肉类具有多重色相特性。以鲜红肉为例,其主色调为红色,但实际光谱反射中还包含其他色相成分。通过光谱反射曲线分析,可精准测定其他色相的存在及其强度。而对于色素形态的检测,光谱反射数据能有效评估色素含量。由于非破坏性检测特性,色度计和分光光度计都是追踪肉类颜色变化的理想工具。需要特别说明的是,不同测色仪器的设计差异显著,这些差异会直接影响测量结果的准确性和可靠性。关于仪器设计的详细讨论,本文不再展开。